Объяснения и примеры
Определение границ экосистем - не слишком сложная проблема при их анализе (гл. 2, разд. 2). До тех пор пока среда на входе и среда на выходе будут рассматриваться как части системы, не имеет значения, как ее ограничить: при помощи естественных ориентиров или произвольно - так, как это удобно. Сложность возникает, однако, при попытке ограничить биотическое сообщество, используя для этого слагающие его видовые популяции. На протяжении последних 50 лет экологи растений вели оживленную дискуссию по поводу того, следует ли рассматривать наземные растительные сообщества как дискретные единицы с определенными границами (Clements, 1905, 1916; Braun-Blanquet, 1932, 1951; Daubenmire, 1966) или же как континуум, поскольку отдельные популяции реагируют на градиенты окружающей среды настолько независимо, что сообщества постепенно переходят друг в друга, и всякое выделение дискретных единиц произвольно (Gleason, 1926; Curtis, McIntosh, 1951; Whittaker, 1951; Goodall, 1963 и др.). Уиттэкер (Whittaker, 1967) иллюстрирует эти различные точки зрения следующим примером. Если бы в разгар осеннего разноцветья в Национальном парке Грейт-Смоки-Маунтинс нам предложили выбрать удобную точку, с которой можно видеть весь градиент высот - от долины до гребней хребтов, то мы бы заметили пять по-разному окрашенных зон: 1) многоцветный лес на площадках среди скал; 2) темно-зеленый лес из тсуги; 3) темно-красный дубовый лес; 4) красновато-коричневые заросли из дуба и вереска и 5) светло-зеленый сосновый бор на гребнях. Эти пять зон можно рассматривать как дискретные типы сообществ, а можно считать их частями одного континуума, которые подлежат какой-либо форме градиентного анализа; при этом выявятся распределение популяций отдельных видов и их реакция на изменение условий внешней среды в соответствии с градиентом. Эту ситуацию иллюстрирует рис. 7.24, на котором показано частотное распределение (в виде гипотетических колоколообразных кривых) 15 видов доминирующих деревьев (от а до n), перекрывающихся вдоль градиента, а также 5 до некоторой степени условных типов сообществ (А, Б, В1, В2 и Г), выделение которых основано на пиках одного или нескольких доминантов. Можно привести много доводов в пользу того, чтобы рассматривать склон в целом как одно большое сообщество, поскольку все эти леса связаны друг с другом обменом биогенными элементами, энергией и животными в одну экосистему водосборного бассейна. Как подчеркивалось в гл. 2, такая экосистема представляет собой минимальную единицу экосистемы, поддающуюся функциональному исследованию и регулированию со стороны человека. Однако лесникам или землеустроителям удобнее рассматривать каждую зону как отдельное сообщество, поскольку каждая из них отличается по скорости прироста древесины, качеству древесины, рекреационной ценности, восприимчивости к пожарам и болезням и по многим другим показателям.
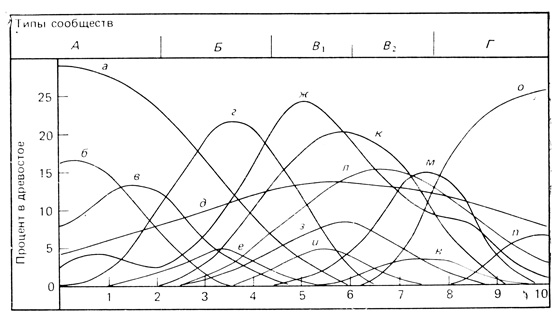
Рис. 7.24. Распределение популяций доминирующих видов деревьев вдоль гипотетического градиента (от 0 до 10), иллюстрирующее размещение популяций в сообществе типа континуум. Кривые распределения каждого вида имеют колоколообразную форму, причем пики относительного обилия (в процентах от общего числа деревьев) приходятся на разные точки вдоль градиента. Некоторые виды устойчивы к более широкому диапазону внешних условий (степень доминирования обычно меньше) по сравнению с другими. Внутри этого большого сообщества можно, используя сочетания двух или более доминирующих видов (виды-индикаторы) и другие признаки, выделить подсообщества (А-Г в верхней части графика). Такое подразделение до некоторой степени условно, но полезно для целей описания и сравнения. Кривые построены по данным ряда исследований распределения видов деревьев по градиенту высоты. (Whittaker, 1954)
Как это часто бывает, концепции и подходы зависят от географии; так, экологи, работающие в областях с постепенно изменяющимися условиями среды и однообразными почвами или субстратами, предпочитают концепцию континуума и разные методы ординации (т. е. статистические методы распределения популяций и сообществ по градиентам). Вместе с тем экологи, работающие в областях с резкими изменениями условий среды и неоднородным рельефом, отдают предпочтение зональной концепции. Методы ординации часто требуют от экологов сравнения сходства (или несходства) последовательных проб, взятых вдоль градиента внешней среды. При этом используется в общей форме такой индекс:
| Индекс сходства (S) = | 2C | , |
| А + В |
где А - число видов в пробе А; В - число видов в пробе В и С - число общих видов в обеих пробах. В работе Уиттэкера (Whittaker, 1967) и Мак-Интоша (McIntosh, 1967) рассматриваются методы ординации и другие приемы градиентного анализа.
Билз (Beals, 1969) непосредственно сравнил изменение растительности на крутых и пологих склонах гор в Эфиопии; результаты этой работы приведены на рис. 7.25. На крутом склоне наблюдается большая разрывность, о чем свидетельствуют несколько острых пиков на кривой индекса несходства (1-S, как было показано выше), рассчитанной для каждой пары соседних проб (рис. 7.25, вверху). Кроме того, на крутом склоне в большей степени, чем на пологих, выражена тенденция к внезапному появлению и исчезновению видов. Как показывает рис. 7.25, внизу, распределения доминирующих видов по частоте при постепенном изменении высоты имеют колоколообразную форму нормального распределения (напоминая гипотетические кривые рис. 7.24). При резком же изменении высоты эти кривые характеризуются резким подъемом и спадом и усеченной вершиной. Билз пришел к выводу, что "вдоль крутых склонов сама растительность может образовывать разрывы, несмотря на непрерывность градиента среды, тогда как на пологих склонах этого может и не быть". Обособлению одного сообщества от другого могут способствовать различные типы взаимодействия между популяциями, например: 1) конкурентное исключение; 2) симбиоз между группами зависящих друг от друга видов и 3) коэволюция групп видов (это будет подробно рассмотрено в следующей главе). Такие факторы, как пожары и антибиотики, также могут создавать четкие границы. Бьюэлл (Buell, 1956) описал ситуацию в парке Итаска (шт. Миннесота), где в пределах основного массива кленово-липового леса имеются довольно хорошо отграниченные островки елово-пихтового леса, не связанные с изменениями рельефа местности. В морских донных сообществах на крутых обрывах также возникает четко выраженная зональность, подобная зональности на горных склонах.
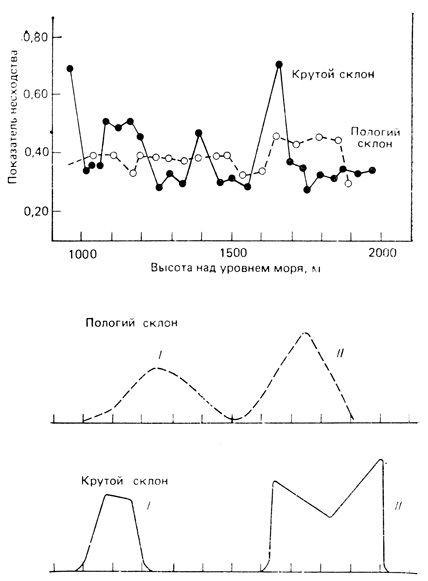
Рис. 7.25. Градиентный анализ изменения растительности на крутом (сплошные линии) и пологом (штриховые линии) склонах одинаковой высоты (Эфиопия). Вверху. График показателей различия между соседними участками вдоль склона. Внизу. Частотное распределение двух видов, Acacia Senegal (I) и Carissa edulis (II). На крутом склоне виды и группы сообществ разграничены более четко. (Beals, 1969)
В тех местах, где происходят резкие изменения по градиенту или где соприкасаются границы двух хорошо отличающихся местообитаний или сообществ, образуется экотон или переходная зона. Часто в этой переходной зоне развивается сообщество, отличающееся от прилежащих сообществ; объясняется это тем, что многим видам нужны в качестве частей местообитаний или мест, где проходят разные стадии жизненного цикла, такие условия, какие имеются в двух или более соприкасающихся и часто сильно различающихся по структуре сообществах. Например, американскому дрозду нужны деревья, на которых он гнездится, и открытые травянистые пространства, где он питается. В хорошо развитые сообщества экотонов входят виды, характерные для каждого из перекрывающихся сообществ, и, кроме того, виды, характерные только для экотона, поэтому число видов и плотность популяций в экотоне выше. Тенденция к увеличению разнообразия и плотности живых организмов на границах сообществ известна под названием краевого эффекта.
В одной из первых работ, где был показан краевой эффект, Бичер (Beecher, 1942) обнаружил, что плотность населения птиц увеличивается с увеличением отношения длины опушки к площади, занимаемой сообществом. Хорошо известно, что плотность певчих птиц выше на территории разного рода усадеб, в окрестностях колледжей и тому подобных местах, которые состоят из смешанных местообитаний и, следовательно, характеризуются большей протяженностью "границ" по сравнению с большими однородными участками леса или полей.
Экотоны могут иметь характерные для них виды, не встречающиеся в соседних сообществах, участвующих в образовании этих экотонов. Например, при изучении населения птиц вдоль градиента развития сообщества (см. табл. 8.2) пробные площадки были выбраны с расчетом, чтобы свести к минимуму влияние контактов с другими сообществами. В некоторых из этих стадий развития сообщества плотность 30 видов птиц составляла не менее пяти пар на 40 га. Однако известно еще около 20 видов птиц, которые обычно гнездятся в сообществах внутренних областей этого региона в целом. Семь из них были встречены в небольшом числе, а остальные тринадцать видов вообще не были зарегистрированы на обследованной однородной территории, в том числе такие обычные виды, как дрозд, синяя птица, многоголосый пересмешник, овсянка индиговая, воробьиная овсянка и садовая иволга. Многим из них деревья необходимы как места для устройства гнезд или в качестве наблюдательных постов, питаются же они преимущественно на земле, в траве или на других открытых участках. Поэтому их требования к местообитаниям лучше всего удовлетворяются в экотонах между лесными и луговыми или кустарниковыми сообществами, а не в местах, занятых лишь одним из этих сообществ. Таким образом, в рассмотренном случае около 40% обычных видов птиц (20 из 50), размножающихся в этом районе, можно считать в основном или полностью экотонными.
Дэвид Пэттон (David Patton, 1975) предложил один из вариантов индекса разнообразия для сравнения отношения протяженности границ к площади в разных ситуациях. Поскольку наименьшее отношение к площади характерно для окружности, именно это отношение принимается за единицу. Исходя из этого, краевой индекс (EI) можно вычислить следующим образом:
| EI = | TP | , |
| 2 ⋅ Aπ |
где ТР - общий периметр площади плюс длина всех линейных границ внутри этой площади, А - площадь и π = 3,14. Квадратная площадка с одним типом растительности характеризуется индексом 1,13. Если на этой площадке четыре разных типа растительности, занимающие одинаковые площади, то дополнительные внутренние границы увеличат значение индекса до 1,69. Если два из четырех типов растительности разделить еще дополнительной границей, то индекс возрастет до 1,97. Два других варианта краевого индекса рассматриваются в работе Тейлора (Taylor, 1977).
Одним из обычных типов экотонов, наиболее важных для человека, является опушка леса. Опушку можно определить как экотон между лесным и травянистым или кустарниковым сообществами. Где бы ни поселился человек, он стремится сохранить поблизости от своих жилищ сообщества опушек. Так, если он селится в лесу, он сокращает лес до отдельных небольших участков, перемежающихся лугами, сельскохозяйственными угодьями и другими, более открытыми местообитаниями. В работе Хокинса (Hawkins, 1940) приводятся карты, на которых зарегистрированы изменения, происшедшие в шт. Висконсин за столетие после появления здесь в 1838 г. первых поселенцев. Если человек селится на открытом месте, то он сажает деревья, создавая подобную же мозаичную структуру ландшафта. Можно сказать, что предпочитаемое местообитание Homo sapiens - это опушка леса, поскольку он не прочь понежиться в тени деревьев и кустарников, но "добывает" себе пищу на лугах и полях. Некоторые виды, обычные для лесов и степей, способны жить на созданных человеком опушках. Другие же, хорошо адаптировавшиеся к жизни на опушках, особенно многие виды трав, насекомых, птиц и млекопитающих, в таких условиях часто увеличиваются в числе и расширяют свои границы обитания по мере создания человеком новых местообитаний типа опушек.
Вообще говоря, охотничье-промысловые виды животных, такие, как олени, кролики, куропатки, фазаны и т. д., предпочитают держаться на опушках. Поэтому человек искусственно создает опушки, сажая съедобные для этих видов растения или растения, дающие им приют и укрытие. Пятнистые местообитания создаются также путем расчисток, выжигания (см. рис. 5.25) и других мероприятий.
Олдо Леополд, которому приписывают честь введения понятия краевого эффекта, писал в своей новаторской работе по охотничьему хозяйству (Leopold, 1933), что "дикая природа - это феномен опушек". Не так давно Хэнссон (Hansson, 1979) рассмотрел значение гетерогенности ландшафта в жизни северных теплокровных животных, которые активны круглый год. Сельскохозяйственные и другие нарушенные пространства предоставляют этим животным зимой больше пищи, чем зрелые ненарушенные леса. Весной и летом, однако, эти последние предоставляют больше пищи.
Не следует думать, что увеличение плотности в экотонах - явление универсальное. Многие организмы ведут себя противоположным образом. Так, не вызывает сомнений, что плотность деревьев в экотоне лесной опушки ниже, чем в лесу. Вырубка на обширных пространствах дождевого тропического леса почти наверняка уменьшает видовое разнообразие и вызывает вымирание многих видов, адаптированных к большим площадям однородного местообитания. Возникает предположение, что наибольшее значение экотоны приобретают там, где люди на протяжении многих столетий сильно модифицировали природные сообщества и осваивали ландшафты, предоставив развитию адаптаций эволюционное время. Например, в Европе, где от лесов остались лишь узкие полоски, дрозды и другие лесные птицы охотнее живут в городах и пригородах, чем родственные им виды в Северной Америке. Но, конечно же, многие другие европейские виды не адаптировались и стали редкими или вымерли.
Как и в большинстве случаев положительных реакций или выгоды, горбатая кривая субсидия - стресс (см. рис. 3.5) применима и к соотношению граница - разнообразие. Чрезмерное увеличение границ (много мелких блоков местообитания) вызывает перегиб кривой разнообразия в сторону снижения. Хотя увеличение границ часто повышает разнообразие, уменьшение площади местообитания снижает разнообразие (тенденция разнообразие - площадь). Теоретический максимум видового бета-разнообразия должен находиться там, где велики или достаточно велики блоки местообитания и велика также общая протяженность границ в регионе (Thomas et al., 1979). Эти противоположные тенденции нужно учитывать при ведении лесного и охотничьего хозяйства и общем планировании ландшафтов. Островная теория биогеографии (будет обсуждаться в следующей главе) может до некоторой степени помочь в определении величины блоков, необходимой для сохранения как отдельных видов, так и разнообразия в целом.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'