Объяснения
Согласно Коуди (Cody, 1966), распределение или подразделение (Allocation) энергии между основными видами активности организма отражает балансы между преимуществами и стоимостью каждого вида активности, которые обеспечивают изменения rmax - внутренней (генетически детерминированной) скорости роста, повышающие в будущем выживание или приспособленность. Прежде всего оплачивается, конечно, выживание и поддержание особи (компонент дыхания) с необходимым добавлением в виде чистой энергии, ассигнованной на рост и размножение (компонент продукции). Крупные организмы (как и большие города) должны направлять большую часть метаболизированной энергии на вход блока поддержания; у мелких организмов эта доля меньше, так как они не обладают большими структурами, требующими поддержания. Естественный отбор, этот бескомпромиссный мастер навязывать функции, требует, чтобы все организмы нашли оптимальное равновесие между расходами энергии на выживание в будущем и расходами на выживание в настоящем.
На рис. 6.23 показаны четыре гипотетических способа распределения энергии между тремя основными статьями расхода: 1) энергия для оплаты конкуренции с другими видами за общие ресурсы; 2) энергия, оплачивающая возможность быть не съеденным хищником (или травоядным, если речь идет о растительном организме); и 3) энергия для производства потомства. Если ущерб от конкуренции и выедания невелик, то большая часть потока энергии может быть направлена на размножение и продуцирование молоди, как показано в случае А. Напротив, избегание конкуренции или выедания может потребовать больше энергии, как показано в случаях Б и В. В случае Г энергия примерно поровну распределяется между этими тремя типами активности. А, Б, В и Г могут обозначать четыре разных вида, у которых давление отбора приводит к такому характеру подразделения энергии, или четыре разных сообщества, в которых такое подразделение присуще большинству видов, или же, как это будет показано в гл. 8, А - обычная ситуация на стадиях колонизации или начальных стадиях сукцессии, когда преобладает r-отбор, тогда как В и Г - ситуация на более зрелых стадиях, когда преобладает K-отбор.
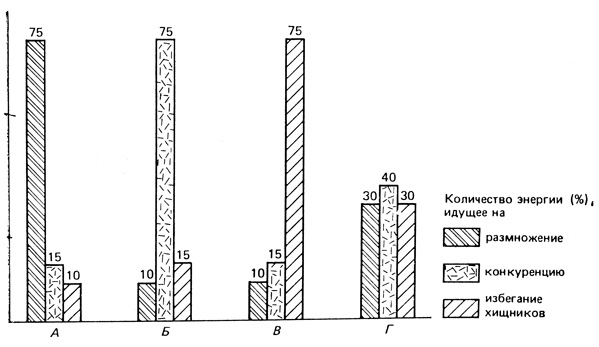
Рис. 6.23. Гипотетическое распределение энергии между тремя основными видами жизнедеятельности, которое необходимо для выживания в четырех резко различающихся ситуациях, когда относительное значение каждого вида жизнедеятельности меняется. (Из Cody, 1966, с изменениями)
Шенер (Schoener, 1971), Коуди (Cody, 1974), Пайк и др. (Pyke et al., 1977), обобщив методы анализа путей распределения и оптимизации энергии, высказали предположение, что в данном случае можно пользоваться методами, аналогичными применяемым при анализе прибыли и затрат в экономике, причем прибыль в экологии - это повышение приспособленности, а затраты - энергия и время, необходимые для обеспечения размножения. Хищник, например, под давлением отбора вынужден увеличивать соотношение между чистой энергией (ассимилированная энергия минус затраты на поимку жертвы) и временем, необходимым для поиска, преследования и поедания добычи. Теоретически увеличить количество энергии, используемой для размножения, можно, либо 1) выбирая более крупную или более питательную жертву или жертву, которую легче поймать, либо 2) уменьшая время и усилия, направленные на поиск и преследование.
Для того чтобы определить оптимальное распределение энергии для данного вида, популяции или данной ситуации в местообитании, было предложено несколько подходов. Один из подходов состоит в том, чтобы выявить, каким образом экологически эквивалентные виды используют морфологические, метаболические и поведенческие адаптации для сохранения приспособленности в сходных местообитаниях, но в различных географических областях. Если параметры приспособленности очень сходны, т. е. наблюдается конвергенция, то можно заключить, что отбор привел к оптимальному решению, несмотря на различия в истории, масштабе времени и генетическом происхождении. Примером служит проведенное Коуди (Cody, 1974) сравнение степных птиц Канзаса и Чили, результаты которого представлены в табл. 7.4.
Другой, более специальный подход состоит в графическом "анализе стратегий" (Levins, 1968), как это показано на рис. 6.24. На рисунке приведены две модели стратегий добывания пищи гипотетическим видом, перед которым стоит задача выбора либо числа пищевых объектов из шести возможных (А), либо определенного числа изолированных участков ("пятен") для добывания пищи (Б). Если особи этого вида питаются каким-то одним пищевым объектом из многих возможных, то им потребуется затрачивать больше усилий на отыскание каждой жертвы, чем если бы они питались любым из шести объектов (кривая ΔS на рис. 6.24, А). Но при увеличении числа объектов питания растут затраты на преследование (одни объекты трудно поймать, другие слишком малы) (кривая ΔР). Оптимальное соотношение дохода и затрат достигается в точке пересечения восходящей и нисходящей кривых, в нашем гипотетическом случае - при питании на четырех объектах (рис. 6.24, А). Взаимодействия с другими видами или другими факторами среды могут сдвинуть оптимум в любом направлении. Конкуренция с другими видами может привести наше гипотетическое животное к "специализации" на питании только одним объектом, если это дает ему преимущество в конкуренции. Или же такой отбор может давать преимущество, когда пищи много. Тогда условия будут диктовать, что лучшая стратегия - это стратегия "генералиста". Пример смещения пищевой избирательности будет приведен ниже.
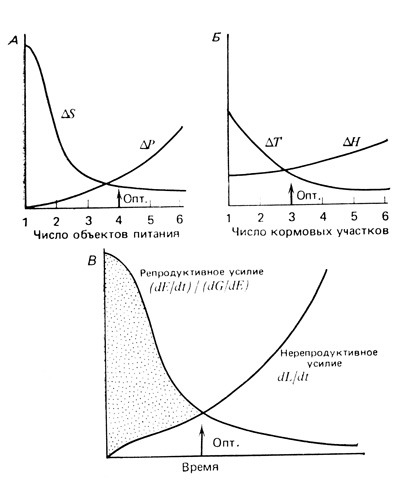
Рис. 6.24. Модели оптимизации стоимости и доходов. Объяснения см. в тексте
Из рис. 6.24, Б видно, что охотничье усилие ΔН возрастает по мере увеличения числа используемых участков для поиска пищи, но это уравновешивается уменьшением времени перехода ΔТ для поимки одного объекта. И вновь оптимум представляет собой компромисс между противоположными тенденциями; в описываемой ситуации это три изолированных охотничьих участка.
Более общая модель приведена на рис. 6.24, В. Здесь по мере увеличения времени, затрачиваемого на добывание энергии (пищи), репродуктивный выход единицы энергии снижается, а нерепродуктивный - растет (в обоих случаях монотонно). Область максимального репродуктивного выхода на рисунке покрыта точками, а оптимальное время питания, обеспечивающее благоприятный баланс между двумя необходимыми путями расхода энергии, вновь соответствует пересечению кривых.
Как отмечалось в разд. "Определения", виды с высоким биотическим потенциалом (r) отбираются обычно в ненасыщенной или неопределенной среде, подверженной периодическим стрессам (таким, как штормы, засухи и т. д.). Виды, у которых энергия распределяется в пользу поддержания и повышения конкурентной способности, чувствуют себя лучше при плотности, равной K (насыщение), или при стабильных физических условиях (мала вероятность сильных неблагоприятных возмущений). По другому пути идут виды, рост популяций которых описывается J-образной кривой. Это хорошие пионерные виды, которые могут быстро использовать нетронутые или недавно накопленные ресурсы, они упруго реагируют на различного рода нарушения. Медленно растущие виды и популяции лучше адаптированы к зрелым сообществам и более устойчивы, но менее упруго реагируют на нарушения (вспомните обсуждение резистентной и упругой устойчивости в гл. 2, разд. 6).
Общая модель r- и K-отбора, предложенная Мак-Артуром (MacArthur, 1972), показана на рис. 6.25. На рисунке Х1 и Х2 обозначают два конкурирующих генетических аллеля, однако они могут также обозначать виды. В области А (слева от точки С), где плотность низка, а пищи (или света и биогенных элементов в случае растений) много, побеждает быстрее размножающийся вид или аллель Х2 (dX/Xdt - удельная скорость роста, как описано в разд. 2), т. е. мы имеем дело с r-отбором. В области В (справа от точки С) вид Х1 растет быстрее, чем Х2, и поэтому побеждает, т. е. имеет место K-отбор. Мак-Артур отмечает, что K-отбор преобладает в тропиках с относительно слабо выраженной сезонностью, тогда как r-отбор преобладает в умеренных зонах, где сезонность хорошо выражена; в таких условиях численность популяции растет экспоненциально, а затем следует ее катастрофический спад.
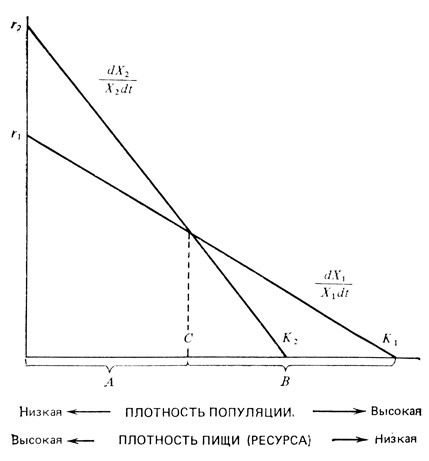
Рис. 6.25. Модель r- и K-отбора Мак-Артура (MacArthur, 1972). Скорости размножения двух аллелей (или видов) изображены в виде функций плотности популяции и ресурса. Когда плотность превышает С, а ресурсов мало, X1 растет быстрее, чем Х2, но Х2 растет быстрее и 'победит' в естественном отборе, если плотность ниже С. В соответствии с этим А представляет собой область r-отбора, а В - область K-отбора
На величину выводка у птиц (число яиц или молоди за репродуктивный период) влияют не только смертность и выживание, она зависит также от r- и K-отбора (см. Lack, 1966, 1968; Cody, 1966; Hutchinson, 1975, гл. 3). У оппортунистических видов выводки больше, чем у сбалансированных, что видно при сравнении птиц умеренной зоны и тропиков.
Обозначение вида как r- или K-стратега может быть ошибочным из-за чрезмерного упрощения классификации, поскольку многие популяции изменчивы в этом смысле или занимают промежуточное положение. Однако Пианка (Pianka, 1970) считает, что в природе наблюдается бимодальность относительна r- и K-отбора, связанная с размерами тела и временем генерации. Он утверждает, что любая из стратегий (либо r, либо K) обладает преимуществом по сравнению с любым компромиссом. Шеффер (Schaeffer, 1976) попытался уточнить эту концепцию, оценивая преимущества и недостатки размножения на разных стадиях и в разное время жизненного цикла.
Левине (Levins, 1968) в книге "Эволюция в меняющейся среде" пришел к заключению, что специализация в эволюции видов ограничивается неопределенностью среды обитания. Для отбора в нестабильных условиях лучше быть "генералистом" и обладать высокой rmax. Сообщества в таких условиях могут быть только очень слабо организованными. Специализация и организация могут подняться до высокого уровня лишь тогда, когда непредсказуемость среды мала (согласно Левинсу). До каких пределов могут совместно действующие группы популяции и сообществ снизить вариабельность среды и открыть тем самым путь для формирования организации более высокого уровня, как это иногда (но не всегда) происходит в человеческом обществе? Этот вопрос остается пока без ответа.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'