Примеры
Аспекты распределения энергии в связи с трофическими уровнями, размером организма, наличием или отсутствием терморегуляции обсуждались в гл. 3, разд. 4 (см., в особенности, модель универсального потока энергии на рис. 3.12). Более частный случай представлен в табл. 6.6, где сравниваются пути распределения ассимилированной энергии (А на рис. 3.12) между продукцией Р (рост и размножение) и дыханием R (поддержание) у четырех видов, представляющих хищников и растительноядных, а также позвоночных и членистоногих. В общем хищники (болотные крапивники и пауки-охотники) тратят больше ассимилированной энергии на поддержание (добывание пищи, защита территории и т. д.), чем растительноядные (хлопковые крысы и гусеницы). Сходным образом крупные эндотермные животные (теплокровные позвоночные) в отличие от мелких экзотермных (членистоногих) направляют большую часть ассимилированной энергии на дыхание. Мак-Нейл и Лоутон (McNeil, Lawton, 1970) отмечают, что у водных видов стоимость дыхания выше, чем у наземных видов, однако на вопрос о том, является ли это различие основным и почему так должно быть, ответа до сих пор не получено.

Таблица 6.6. Распределение ассимилированной энергии между продукцией (рост и размножение) и дыханием (поддержание)
Интересно сравнить распределение энергии у пауков-охотников и тенетников. Поскольку паутина содержит много белка, на синтез шелка затрачивается много энергии, но многие науки используют шелк повторно, съедая старую паутину при постройке новой. Таким образом, цена паутины снижается. Пиколл и Витт (Peacall, Witt, 1976) установили, что на образование шелка у одного из пауков, который реутилизирует свою паутину, тратится примерно четвертая часть того количества калорий, которые израсходованы на работу по строительству паутины и ее регулярному ремонту. Общая энергетическая стоимость паутины составляет около половины основного потребления энергии, и это меньше, чем затраты на охоту у некоторых не строящих паутины видов. Возможно, это урок для человека: вид, который конструирует дорогостоящие и сложные механизмы, может удешевить их путем повторного использования материалов.
Теория, согласно которой хищник оптимизирует энергетику дохода и затрат, выбирая наиболее благоприятный размер жертвы в зависимости от общего обилия пищи, была экспериментально проверена и подтверждена Вернером и Холлом (Werner, Hall, 1974). Эти исследователи предлагали ушастым окуням в разных комбинациях ветвистоусых рачков разной величины и регистрировали размеры избираемой окунями жертвы. Когда абсолютное количество пищи было невелико, съедались подряд жертвы всех размеров. При возрастании количества пищи рыбы выедали наиболее крупных рачков, не трогая представителей мелких размерных классов. Таким образом, при увеличении количества пищи (и наоборот, когда пищи становится мало) рыбы способны менять стратегию, становясь из "генералистов" "специалистами".
Для иллюстрации r- и K-отбора приведем два примера: амброзию, которая растет по залежам и другим недавно нарушенным местообитаниям, и травянистое растение Dentaria laclniata, обитающее в относительно стабильном нижнем ярусе леса. Эти растения сравниваются по продукции семян и максимальному репродуктивному усилию в табл. 6.7. Амброзия продуцирует примерно в 50 раз больше семян, чем Dentaria, и тратит значительно большую долю чистой энергии на размножение.

Таблица 6.7. Противоположные стратегии размножения двух травянистых растений, а также полевого и лесного сообществ, в которых они растут: пример r- и K-отбора (Newell, Tramer, 1978)
Золотарник дает пример целого ряда репродуктивных стратегий, промежуточных между крайними стратегиями r- и K-отбора. На рис. 6.26 приведены данные, характеризующие репродуктивное усилие и накопление биомассы для шести популяций четырех видов золотарника рода Solidago. У популяции вида 1, который растет на сухих открытых полях и нарушенных местообитаниях, поддерживающая биомасса листьев мала и около 45% чистой продукции направлено на образование репродуктивных тканей. Популяция 6, встречающаяся во влажных лиственных лесах, напротив, вкладывает большую часть энергии в листья и только 5% затрачивает на размножение. Другие популяции встречаются в промежуточных по влажности и стабильности местообитаниях и в соответствии с этим распределяют энергию в промежуточных пропорциях.
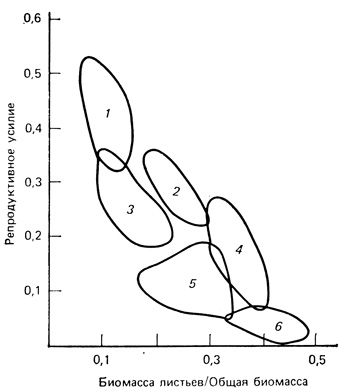
Рис. 6.26. Репродуктивное усилие (отношение сухой массы репродуктивных тканей к общей сухой массе надземных тканей) как функция отношения массы листьев к общей массе в шести популяциях четырех видов золотарника (Solidago). Популяция 1 принадлежит виду, который растет на сухих открытых полях или нарушенных почвах; популяция 6 встречается во влажных лиственных лесах, остальные популяции занимают промежуточные по влажности и стабильности местообитания. (По Abrahainson, Gadgil, 1973)
Солбриг (Solbrig, 1971) описал случай, когда и r-, и K-стратеги встречаются у одного и того же вида. У обычного одуванчика (Taraxacum officinale) имеется несколько линий, или вариететов, различающихся по генотипам, контролирующим распределение энергии. Растения одной линии первыми занимают нарушенные местообитания и продуцируют много мелких рано созревающих семян. Растения другой линии встречаются в менее нарушенных местообитаниях, больше энергии направляют в листья и стебли и продуцируют относительно меньше поздно созревающих семян. Если обе линии произрастают на хорошей почве, то более плодовитая линия оказывается затененной менее плодовитой. Таким образом, первая линия лучше приспособлена для заселения новых местообитаний и считается r-стратегом; вторая линия характеризуется лучшей конкурентоспособностью, и ее следует считать K-стратегом.
Хотя вариабельность и нарушение местообитаний должны благоприятствовать r-стратегам, это не значит, что K-стратеги исчезают.
В адаптированных к пожарам сообществах, например в сообществах калифорнийского чапараля (гл. 5, разд. 5), растения запасающие большое количество энергии в подземных частях и размножающиеся побегами, приспособлены к переживанию периодических пожаров так же хорошо, как растения, вкладывающие свое будущее в семена (см. Carpenter, Recher, 1979).
Другие примеры распределения энергии в растительном царстве приведены в работах Харпера (Harper, 1977) и Гэджила и Солбрига (Gadgil, Solbrig, 1972).
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'