Примеры
Галапагосские вьюрки представляют собой классический пример аллопатрического видообразования (вследствие географической изоляции) с последующим смещением признаков. Этот случай был впервые описан Дарвином, который посетил Галапагосские острова во время своего знаменитого плавания на "Бигле". В условиях изоляции на разных островах от общего предка возникла целая группа видов; они претерпели столь сильную адаптивную радиацию, что при повторных заселениях различных островов заполняли самые разнообразные потенциальные ниши. Среди современных видов есть тонкоклювые насекомоядные, толстоклювые зерноядные, питающиеся на земле и на деревьях, крупные и мелкие вьюрки и даже один вьюрок, похожий на дятла, хотя он вряд ли выдержал бы конкуренцию с настоящим дятлом и может выжить лишь в отсутствие этих птиц (см. Lack, 1947). На рис. 8.12 показаны вариации размера клюва одного из галапагосских вьюрков в зависимости от того, обитает ли он на острове один или же вместе с двумя близкородственными видами на другом, более крупном острове. В этой ситуации (Б на рис. 8.12) происходит "смещение" размера клюва в направлении увеличения его толщины, так что он не перекрывается по размерам с клювами двух конкурентов. В результате конкуренция за пищу уменьшается, так как каждый из трех видов адаптирован к питанию семенами разной величины.
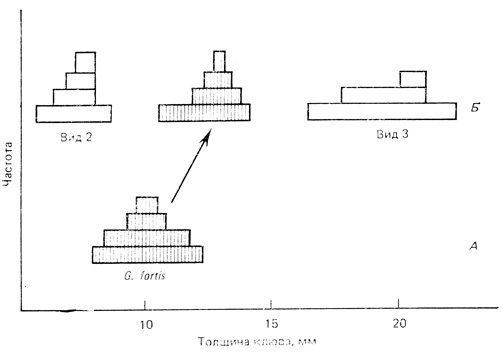
Рис. 8.12. Величина клюва у Дарвинова вьюрка Geospiza fortis, живущего на одном из Галапагосских островов в отсутствие конкурентов (А) и на других островах вместе с конкурентами - другими видами Geospiza (Б). Увеличение размеров клюва в присутствии конкурентов - пример смещения признаков. (Lack, 1947)
Засоленные марши Британских островов дают нам пример симпатрического видообразования в результате гибридизации и полиплоидии. Когда Spartina alterniflora, злак, произрастающий на засоленных маршах Америки, попал на Британские острова, он стал скрещиваться с местным видом Spartina maritima. Возник новый полиплоидный вид Spartina townsendii, который заселил теперь голые, прежде илистые отмели, где не росли местные виды.
Другой пример быстрого естественного отбора, вызванного вмешательством человека, - это так называемый "индустриальный меланизм": возникновение темной окраски у бабочек в промышленных районах Англии, где стволы деревьев заметно потемнели от копоти, которая погубила лишайники, придававшие стволам светлый оттенок. Кеттлуэлл (Kettlewell, 1956) экспериментально показал, что темные бабочки лучше выживают на темных (закопченных) стволах, а светлые - на обычных; вероятно, это вызвано тем, что птицы выборочно истребляют особей, не имеющих покровительственной окраски. Этот пример, естественно, приводит нас к проблеме прямого, или целенаправленного, отбора, осуществляемого человеком.
Искусственный отбор: доместикация. Отбор, который производит человек с целью приспособить растения и животных к своим нуждам, называют искусственным отбором. Доместикация* растений и животных - это не только изменение генетики вида, так как при этом необходима взаимная адаптация одомашниваемого вида и доместикатора. Доместикация, следовательно, ведет к особой форме мутуализма. Будучи эгоцентричным, человек впадает в заблуждение и считает, что, одомашнивая другой организм путем искусственного отбора, он просто "покоряет" природу, приспособляя ее для наших целей. На самом же деле доместикация вызывает у человека такие же изменения (если не генетические, то, во всяком случае, экологические и социальные), как и у одомашненных видов. Поэтому человек в той же мере зависит от сельскохозяйственных растений, как и сельскохозяйственные растения зависят от нас. Общество, хозяйство которого зависит от кукурузы, развивается в культурном отношении совершенно по-иному, нежели общество, занятое пастбищным скотоводством. Еще вопрос: кто у кого в рабстве? Этот же вопрос можно отнести к взаимоотношениям человека и его машин. Взаимоотношения человека с трактором, по-видимому, не слишком отличаются от взаимоотношений человека с лошадью; разница только в том, что трактор требует гораздо больше высококачественной энергии (горючего) и выделяет ядовитые отходы!
* (Для искусственного отбора растений многие предпочитают термин "окультуривание". Здесь мы используем термин "доместикация" (одомашнивание) в общем смысле для растений и для животных. - Прим. автора.)
Хорошим примером взаимозависимости между одомашниваемым видом и доместикатором-человеком служит искусственный отбор зерновых - основа "зеленой революции". Повышение урожайности получено путем отбора на большее отношение зерна (или других съедобных частей) к поддерживающим тканям (листья, корни, стебли и т. д.), иными словами, повышению урожайности выше определенного уровня приносится в жертву адаптивная способность растения к самоподдержанию. Таким образом, высокоинбредные линии требуют мощных вкладов энергии, удобрений и пестицидов, что вызывает глубокие изменения в социальной, экономической и политической структуре общества. Для многих отсталых стран эти требования к социально-экономическому укладу и ресурсам являются наибольшим препятствием для использования высокоурожайных сортов, необходимых для повышения производства продуктов питания.
Некоторые из наиболее серьезных проблем, стоящих перед человеком, связаны с одомашненными растениями и животными, которые "вырвались из плена" (т. е. одичали) и, вернувшись в природу, превратились в настоящий бич. Одичавший организм отличается от своего дикого предка тем, что он перенес период искусственного отбора, приобретя некоторые новые свойства и утратив некоторые "дикие". Вернувшись к жизни в природе, одичавший организм вновь подпадает под действие естественного отбора, который благоприятствует свойствам, необходимым для самостоятельного выживания. Например, отбор работает против пятнистой или светлой окраски и крупных размеров домашних свиней, если они попадают в дикую природу. Поэтому одичавшие свиньи становятся более мелкими и темными. Сочетание искусственного и естественного отбора приводит к тому, что такие растения и животные процветают в частично измененных или нарушенных местообитаниях. Например, одичавшие свиньи и козы могут произвести опустошительные разрушения растительности островов или районов, из которых изъяты крупные хищники.
Островная биогеография. Острова представляют собой природные лаборатории для изучения эволюции, зачаровывая биологов и экологов со времени посещения Дарвином Галапагосских островов. Сложная взаимосвязь изоляции, естественного отбора и видообразования привлекает внимание и сейчас, особенно после того, как Мак-Артур и Уилсон (MacArthur, Wilson, 1963, 1967) опубликовали свою теорию островной биогеографии. В простейшей формулировке эта теория состоит в том, что число видов на острове определяется равновесием между иммиграцией новых видов и вымиранием уже существующих. Поскольку число видов увеличивается и уменьшается примерно логарифмически и поскольку скорости иммиграции и вымирания зависят от величины острова и расстояния до материка, служащего источником видов, общая модель равновесия может быть изображена графически так, как это сделано на рис. 8.13. Показаны четыре точки равновесия, соответствующие 1) небольшому удаленному острову, на котором предсказывается существование немногих видов; 2) большому, близкому к материку острову, который должен быть населен большим числом видов; 3) большому удаленному острову и 4) небольшому близкому острову, которые занимают промежуточное положение в отношении видового богатства. Теория равновесия была проверена Симберлоффом и Уилсоном (Simberloff, Wilson, 1969), которые уничтожили всех членистоногих (путем обработки инсектицидом) на одном из небольших мангровых островов из группы Флорида-Кис и наблюдали реколонизацию. В обзоре Симберлоффа (Simberloff, 1974) содержатся более подробные сведения об островной биогеографии.
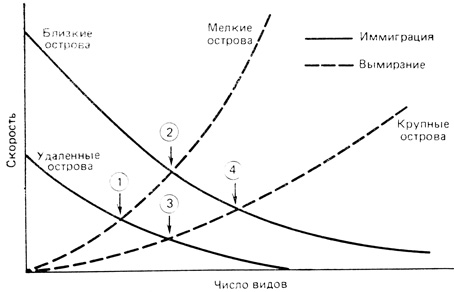
Рис. 8.13. Теория островной биогеографии. Число видов на острове определяется равновесием между скоростями иммиграции и вымирания. Показаны четыре точки равновесия, соответствующие разным комбинациям размеров острова и удаления его от материка. (MacArthur, Wilson, 1963)
Уилсон и Уиллис (Wilson, Willis, 1975), Даймонд и Мей (Diamond, May, 1981) и другие исследователи предположили, что теория островной биогеографии дает основу для планирования резерватов, предназначенных для сохранения природного разнообразия и (или) находящихся под угрозой исчезновения видов. В соответствии с этой теорией один большой резерват предпочтительнее группы мелких равной площади. Если приходится разбивать небольшие парки, то они должны быть расположены близко друг от друга или должны быть соединены коридорами, облегчающими расселение животных. Резерват в форме круга, когда максимизируется отношение площади к периметру, предпочтительнее вытянутого резервата. Резерваты или парки, находящиеся внутри континентальных областей, редко бывают изолированы в такой степени, как океанические или прибрежные острова, поэтому неясно, применима ли к ним модель Мак-Артура-Уилсона. Некоторые другие проблемы, возникающие при применении островной теории к природным заповедникам, рассмотрены в работах Симберлоффа и Эбиля (Simberloff, Abele, 1976) и Кашлена (Kushlan, 1979).
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'