Объяснения
Характерной чертой структуры сообществ в северных широтах, а также сообществ тропических областей с чередованием дождливых и сухих сезонов является наличие немногих обычных, или доминантных, видов, представленных большим числом особей, и многих редких видов, представленных малым числом особей. Во влажных тропиках, где нет смены сезонов, обычно насчитывается множество видов с низким относительным обилием. Общую тенденцию увеличения числа видов с севера на юг иллюстрирует рис. 7.15. Другая общая тенденция, или закон, природы состоит в том, что число видов возрастает с увеличением площади и, вероятно, также с увеличением эволюционного времени, в течение которого происходили колонизация, специализация ниш и видообразование (см. Preston, 1960; Sanders, 1968).
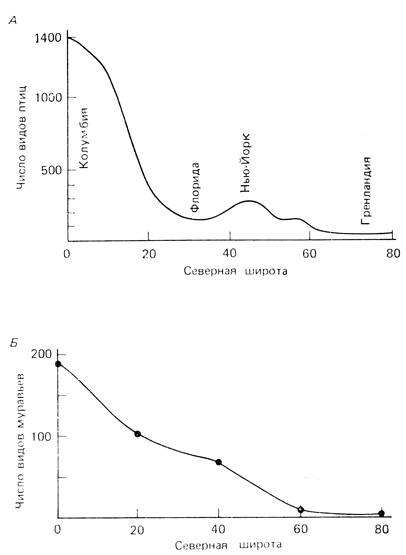
Рис. 7.15. Широтный градиент числа видов гнездящихся наземных птиц (А) и числа видов муравьев (Б). (Fischer, 1960)
Для анализа видового разнообразия в разных ситуациях применяют два широких подхода: 1) сравнения, основанные на формах кривых относительного обилия или доминирования - разнообразия, и 2) сравнения, основанные на индексах разнообразия, представляющих собой отношения или другие математические выражения зависимости между числом видов и их значимостью. Позднее мы более подробно рассмотрим эти подходы, но прежде всего важно уметь различать два основных компонента видового разнообразия, которые могут по-разному реагировать на географические, физические и эволюционные факторы. Один из главных компонентов - видовое богатство (многообразие), или плотность, видов; это просто общее число видов, которое в сравнительных целях обычно выражается как отношение числа видов к площади или числа видов к числу особей. Часто, но далеко не всегда, существует логарифмическая зависимость между числом видов и числом особей (прямая линия в полулогарифмических координатах), и число видов представляет собой степенную функцию от площади (прямая линия в двойных логарифмических координатах), как показано на рис. 7.16, А, Б. Из рис. 7.16, Б видно, что на увеличение числа видов с увеличением площади оказывают влияние как разнообразие местообитаний, так и площадь per se. Соотношения виды/площадь обычно используются для определения оптимальной величины пробы и для предсказания числа видов на большей площади, чем та, где брались пробы. По данным Коннора и Мак-Коя (Connor, McCoy, 1979), хорошее приближение кривых виды/площадь получается при использовании нескольких различных уравнений (в том числе степенной функции в двойной логарифмической шкале), но ни одно из них не имеет биологического смысла.
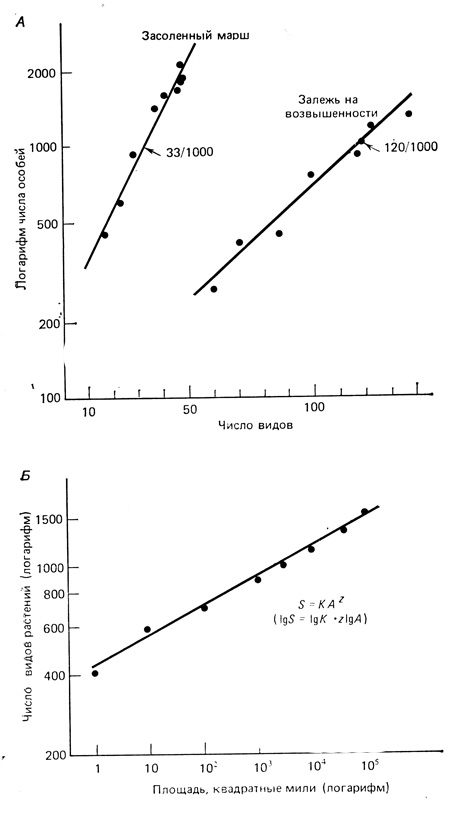
Рис. 7.16. А. Соотношение между числом видов и числом особей на засоленных маршах и залежных землях в Джорджии. При логарифмической зависимости в качестве индекса разнообразия можно использовать число видов на 1000 особей. Б. Зависимость между площадью и числом видов растений (Великобритания). Линии регрессии проходят с наклоном от 0,1 до 0,35. Низкие величины характерны для континентов, высокие - для островов. (См. Connor, McCoy, 1979)
Второй важный аспект разнообразия - выровненностъ относительного распределения особей среди видов. Например, две системы, каждая из которых состоит из десяти видов и 100 особей, могут иметь одинаковые индексы разнообразия S/N, но совершенно различные индексы выровненности в зависимости от распределения 100 особей между десятью видами. Так, в одном крайнем случае распределение может иметь вид: 91-1-1-1-1-1-1-1-1-1 (минимальная выровненностъ и максимальное доминирование), а в другом крайнем случае - по десяти особей на каждый вид (максимальная выровненностъ и отсутствие доминирования). Выровненность, как правило, высока и постоянна среди популяций птиц (вероятно, это обусловлено их территориальным поведением), а различия этого показателя в разных сообществах и географических зонах определяются главным образом видовым богатством (Tramer, 1969). Напротив, у растений и фитопланктона выровненность в среднем низка, и оба компонента подвержены значительным вариациям (Austin, Tomoff, 1978).
Наилучший способ представить оба компонента разнообразия - построить график, где в логарифмическом масштабе отложено число особей (биомасса, продуктивность или другие подходящие меры значимости) каждого вида, а на оси х - ранжированная последовательность видов от наиболее обильного до наименее обильного. Линия, соединяющая точки или проходящая близко от них, названа Уиттэкером (Whittaker, 1965, 1972) кривой доминирования - разнообразия. Еще одно удачное название - "кривая значимости видов" - было предложено Пианкой (Pianka, 1978). На рис. 7.17 показаны три основные формы, которые может иметь такая кривая, построенная для гипотетической выборки из 1000 особей и 20 видов. Если наиболее обильный вид в два раза многочисленнее следующего за ним по обилию, а этот в свою очередь вдвое многочисленнее третьего и т. д., то на графике мы получаем прямую линию (рис. 7.17, I). Отсюда можно предположить, что первый вид занимает половину доступного пространства ниш, второй - половину оставшегося пространства (25% исходного) и т. д. Иными словами, каждый вид занимает прежде всего свободную нишу, не перекрывающуюся с другими. Если, однако, пространство ниш поделено на случайные, соприкасающиеся и неперекрывающиеся участки, то получится совершенно другая кривая (рис. 7.17, II). Эта кривая изображает модель "разломанного стержня", предложенную Мак-Артуром (MacArthur, 1957). Эти две возможности представляют крайние случаи, обычно же распределение в природе имеет вид промежуточной S-образной кривой (рис. 7.17, III), что указывает на более сложный характер дифференциации и перекрывания ниш. Об этом свидетельствует логнормальное распределение видов по значимости (рис. 7.17, III). (См. Preston, 1948; Whittaker, 1972, и рис. 7.21, Б.) У групп с интенсивной межвидовой конкуренцией и территориальным поведением, например у лесных птиц, характер распределения соответствует представлению о неперекрывающихся случайных нишах, как на кривой II; простые геометрические последовательности (кривая I) характерны для некоторых растительных сообществ в суровых условиях окружающей среды. Как было подчеркнуто в разд. 2, большинство видов в природных открытых системах сосуществует в условиях соревнования за ресурсы, а не в условиях прямой конкуренции; множество адаптаций дает возможность делить ниши без конкурентного исключения из местообитания. Таким образом, наиболее вероятную модель изображает кривая III, особенно для ненарушенных сообществ.
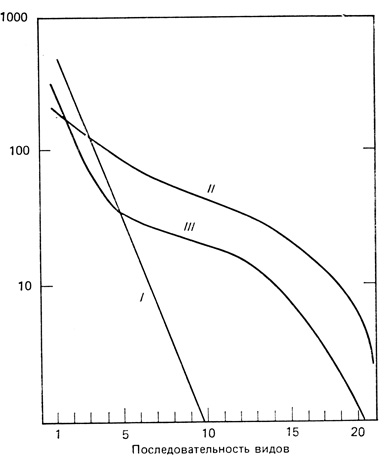
Рис. 7.17. Кривые доминирования - разнообразия для гипотетической выборки, состоящей из 1000 особей, относящихся к 20 видам. По оси ординат отложено число особей каждого вида, а по оси абсцисс - порядковый номер вида в последовательности от более обильного к менее обильному. I - геометрический ряд с приоритетами в захвате ниш; II - ниши не перекрываются, их величина случайна; III - промежуточная система, соответствующая S-образной кривой; ниши многомерны и перекрываются, что обусловливает логнормальное распределение видов по значимости. (Whittaker, 1965)
В общем расположение видов по степени значимости, как на рис. 7.17 и 7.18, не только точно отражает богатство и относительное обилие видов как компонентов видового разнообразия, но и объясняет, каким образом подразделяется пространство ниш. Чем выше кривая и чем более она уплощена, тем больше при данном числе видов общее разнообразие; так, на рис. 7.17 разнообразие II> разнообразие III> разнообразие I. Чем круче идет кривая, тем меньше общее разнообразие и сильнее доминирование одного или нескольких видов. В стрессовых ситуациях независимо от того, вызваны ли они естественными причинами (например, суровыми погодными условиями) или антропогенным воздействием (например, загрязнения), кривая становится более крутой. Таким образом, кривую доминирования - разнообразия можно использовать для оценки влияния нарушений на видовую структуру.
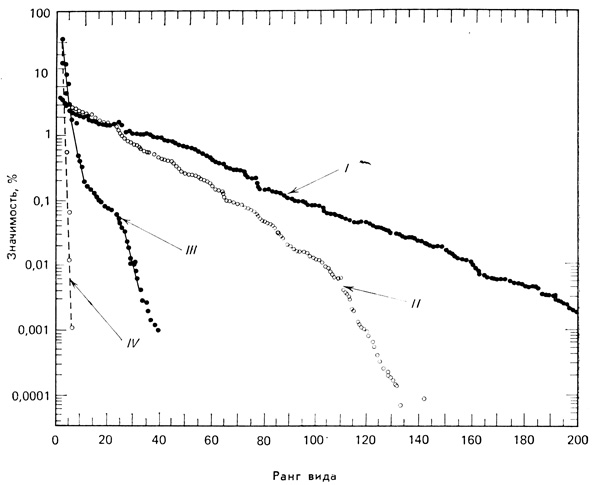
Рис. 7.18. Сравнение кривых доминирования - разнообразия для двух тропических лесов и двух лесов умеренной зоны. Оценки значимости видов в лесах умеренной зоны основаны на чистой годовой продукции, в сухом лесу Коста-Рики - на площади оснований (площадь сечений всех стволов данного вида), в лесу Амазонки - на биомассе надземных частей. (Hubbel, 1979.) I - влажный тропический лес (Манаус, Бразилия); II - сухой тропический лес (Гуанакасте, Коста-Рика); III - сырой лес умеренной зоны (Смоки-Маунтинс, Теннесси); IV - горный (субальпийский) лес умеренной зоны (Смоки-Маунтинс, Теннесси)
При другом подходе используются индексы разнообразия. В табл. 7.5 приведены четыре наиболее употребительных индекса и некоторые их варианты. А - индекс видового богатства; Г - индекс выровненности; Б и В содержат комбинацию обоих компонентов и, таким образом, представляют собой обобщенные индексы разнообразия. Индекс Симпсона показывает "концентрацию" доминирования, так как его величина тем больше, чем сильнее доминирование одного или немногих видов. Единица минус индекс Симпсона (или просто обратная ему величина) становится индексом разнообразия, сравнимым с другими.

Таблица 7.5. Некоторые полезные показатели разнообразия
1 (Simpson, 1949.)
2 (См. Margalef 1958, - логарифмический индекс; Menhinick, 1964, - индекс квадратного корня; Н. T. Odum, Cantlon, Kornicker, 1960, - индекс S/1000 особей.)
3 (Shannon, Weawer 1949; Margalef, 1968.)
4 (См. Pielou, 1966; другие типы показателя выровненности - см. Lloyd, Ghelardi, 1964.)
Примечание. При расчете величин d, H¯ и е обычно пользуются натуральными логарифмами (loge), но для вычисления Н¯ иногда применяют логарифм с основанием 2 (log2), чтобы сразу получить эту величину в битах на особь.
Из двух обобщенных индексов индекс Симпсона придает обычным видам больший вес (поскольку при возведении в квадрат малых отношений ni/N получаются очень малые величины). Индекс Шеннона придает больший вес редким видам. Так как индекс Шеннона заимствован из теории информации и представляет собой формализацию, которая широко используется при оценке сложности и содержания информации в любых типах систем, он лучше всего подходит для целей сравнения в тех случаях, когда нас не интересуют компоненты разнообразия по отдельности. И коль скоро Н¯ вычислено, мы можем, поделив эту величину на логарифм числа видов, легко получить индекс выровненности (Г в табл. 7.5). Индекс Шеннона к тому же почти не зависит от величины пробы и вследствие того, что N - всегда целое число, характеризуется нормальным распределением (Bowman et al., 1970; Hutcheson, 1970); это обстоятельство позволяет использовать обычные статистические методы для проверки значимости различий между средними. Лайонс (Lyons, 1981) показал, что если известно число особей, то в таких статистических тестах могут быть использованы показатели биомассы или продуктивности (нецелые величины), которые с экологической точки зрения часто оказываются более подходящими.
Поскольку стабильные экосистемы, такие, как дождевой лес пли коралловые рифы, обладают высоким видовым разнообразием, возникло искушение сделать вывод, что разнообразие повышает стабильность. Маргалеф (Margalef, 1968) выразил это так: "Эколог видит в разнообразии, измеренном любым способом, проявление возможности построить системы с обратной связью". Однако результаты более поздних исследований и критические обзоры, начиная с симпозиума 1969 г. "Разнообразие и устойчивость экологических систем", материалы которого вышли под редакцией Вудвелла и Смита (Woodwell, Smith, 1969), и кончая статьями в Трудах I международного экологического конгресса под редакцией Ван Доблена и Лоу-Мак-Коннелла (Van Doblen, Lowe-Mc-Connell, 1975), показали, что связь между видовым разнообразием и устойчивостью сложнее, что положительная корреляция может быть вторичной и что, напротив, стабильность экосистемы может обусловливать высокое разнообразие. Хастон (Huston, 1979) пришел к выводу, что экосистемы, названные им "неравновесными", т. е. экосистемы, претерпевающие периодические возмущения, как правило, характеризуются большим разнообразием, чем "равновесные" экосистемы, в которых сильнее выражены доминирование и конкурентное исключение. Вместе с тем Мак-Нотон (McNaughton, 1978) на основе изучения залежей и восточноафриканских злаковников заключил, что на уровне первичных продуцентов (растительности) видовое разнообразие служит средством функциональной стабильности сообщества. Однако, как говорится, хорошенького понемножку; многочисленные виды, вступая в конкурентную борьбу друг с другом, могут дестабилизировать экосистему (см. May, 1973). В природе нигде и никогда не достигается максимальное теоретическое разнообразие, т. е. не бывает так, что одновременно одинаково значимы многие виды; как правило, одни виды всегда более редки, чем другие. В тех случаях, когда разнообразие высоко, в среднем, по-видимому, достигается 80% максимальной выровненности (Odum, 1975). В ходе экологической сукцессии видовое разнообразие обычно увеличивается. Однако это увеличение не всегда затрагивает старые или зрелые стадии, о чем будет сказано в следующей главе (разд. 1 и 2).
Основная трудность при изучении видового разнообразия состоит в том, что исследователи имеют дело только с частями сообществ, обычно с таксономическими группами (например, птицами или насекомыми) или, в лучшем случае, с одним трофическим уровнем. Для оценки разнообразия всего сообщества требуется, чтобы роли разных ниш и организмов разных размеров были "взвешены" каким-то образом на основе общего знаменателя, такого, например, как энергия. Некоторые возможные способы оценки см. Odum, 1982.
Представляется вероятным, что стабильность более тесно связана с функциональным, а не со структурным ("урожай на корню") разнообразием (см. гл. 6, разд. 2).
Для того чтобы разнообразие внутри одного местообитания или типа сообщества не путать с разнообразием ландшафта или региона, который содержит смесь местообитаний, Уиттэкер (Whittaker, 1960) предложил следующие термины: 1) альфа-разнообразие для разнообразия внутри местообитания или внутри сообщества; 2) бета-разнообразие для разнообразия между местообитаниями и 3) гамма-разнообразие для разнообразия в обширных регионах биома, континента, острова и т. д.
Рассматривая трофические уровни, отдельные хорошо изученные зоны, такие, как донное население водоемов, или другие части сообщества, мы обнаруживаем, что большое влияние на видовое разнообразие оказывают функциональные связи между этими уровнями. Например, количество травоядных или хищников сильно влияет на разнообразие травостоя или жертв. Умеренное "хищничество" часто снижает плотность доминантов, предоставляя, таким образом, менее конкурентоспособным видам большие возможности для использования пространства и ресурсов. Разнообразие травянистой растительности на меловых возвышенностях Англии снизилось после того, как были огорожены пастбища для кроликов (Harper, 1969). Вместе с тем чрезмерный выпас оказывает стрессовое воздействие, уменьшая число видов до немногих несъедобных. Пейн (Paine, 1969) обнаружил, что как в тропических, так и в умеренных областях прикрепленные организмы литорали (где пространство в общем более ограничено, чем пища) характеризуются большим видовым разнообразием в тех местах, где активно действуют хищники и первого, и второго порядка. В таких ситуациях искусственное удаление хищников снижает видовое разнообразие всех прикрепленных организмов независимо от того, питались ими хищники или нет. Пейн приходит к выводу, что "локальное видовое разнообразие непосредственно зависит от эффективности, с которой хищники предотвращают монополизацию одним видом основных необходимых ресурсов среды". Этот вывод не всегда обязателен для местообитаний, где конкуренция за пространство менее напряженная. Деятельность человека, как правило, направлена в сторону уменьшения разнообразия и поощрения монокультур, однако часто человек увеличивает разнообразие местообитаний в ландшафте в целом, создавая открытые пространства в лесу, сажая деревья в прерии, интродуцируя новые виды и т. д. Разнообразие растений и мелких певчих птиц намного больше в старых, давно заселенных районах, чем во многих природных местообитаниях (гл. 2, разд. 7).
Виды не всегда служат лучшей экологической единицей для оценки разнообразия, так как различные стадии жизненного цикла или различные жизненные формы одного и того же вида часто занимают разные местообитания и экологические ниши и вносят, таким образом, свой вклад в разнообразие экосистемы. Гусеница и бабочка или лягушка и головастик играют значительно более разнообразные роли в сообществе, чем два вида гусениц или взрослых лягушек. Харпер (Harper, 1977) заметил, что "различные стадии жизненного цикла растений в такой же степени обусловливают разнообразие экосистемы, как и многочисленные виды". Следовательно, для оценки разнообразия не нужно быть опытным систематиком (или нуждаться в его помощи). Например, в коллекции насекомых, собранных сачком, нужно только разобрать особей, у которых есть четкие морфологические и размерные отличия, при этом нет необходимости знать видовые названия или выяснять, принадлежат ли разные "сорта" к одному виду или нет. Иными словами, ni в формулах разнообразия могут быть представлены разными морфологическими типами, которые будут скорее всего представлять разные экологические типы (поскольку, как уже отмечалось, различия в размерах и морфологии указывают на различия ниш).
Рассмотрим генотипическое разнообразие, которое остается скрытым, если характеризовать сообщество только на уровне видов. Как поясняет Вайда (Vida, 1978), классическая теория основывалась на том, что особи в популяции должны быть гомозиготными по аллелям, дающим наибольшую приспособленность. Это видно на приведенной ниже схеме, где некая гипотетическая особь гомозиготна по всем, за исключением одной из шести пар аллелей:
a1b1c1d1e1f1 ____________ ____________ a1b1c1d1e2f1
Альтернативная теория утверждает, что особи гетерозиготны по большинству локусов, и высокий полиморфизм поддерживается разными формами сбалансированного отбора, как показано на следующей схеме, где наблюдается гетерозиготность по всем аллелям, за исключением одного. Разнообразие аллелей здесь намного больше:
a1b5c2d1e1f5 ____________ ____________ a3b2c6d1e2f4
Теория "сбалансированного полиморфизма" представляется более приемлемой, что подтверждается современными биохимическими методами, которые могут выявить скрытую генетическую изменчивость; вероятно также, что полиморфизм поддерживается естественным отбором. В отсутствие такой генотипической изменчивости виды оказались бы неспособными адаптироваться к новым ситуациям и, следовательно, должны были бы вымереть в изменяющейся среде.
Разнообразие видов, стадий жизненных циклов и генетических типов не исчерпывает собой набор элементов, составляющих разнообразие сообщества. Структуры, формирование которых обусловливается распределением организмов в среде или их взаимодействием со средой, рассмотрены Хатчинсоном (Hutchinson, 1953). Различные способы аранжировки одновременно представленных в сообществе организмов характеризуют его структурное разнообразие*. Например:
* (Пиелу (Pielou, 1966а) использует термин "структурное разнообразие" в более узком смысле для обозначения степени разобщения особей одной популяции от особей другой.)
1. Стратификационные принципы (вертикальная слоистость, ярусность растительного покрова и структура почвенных профилей).
2. Зональность (горизонтальная разобщенность, вертикальная поясность в горах или в литоральной зоне).
3. Характер активности (периодичность).
4. Структура пищевой сети (сетевая организация пищевых цепей).
5. Репродуктивные системы (ассоциации родителей и потомства, клоны растений и т. д.).
6. Социальные структуры (стада и табуны).
7. Системы взаимодействия (возникают в результате конкуренции, антибиоза, мутуализма и т. д.).
8. Стохастические структуры (возникают в результате действия случайных сил).
Увеличение разнообразия обусловлено также "краевыми эффектами" - контактами между пятнами контрастирующих типов растительности или физическими местообитаниями. Этот вопрос будет обсуждаться в следующем разделе.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'