Примеры
Результаты одного из оригинальных экспериментов Гаузе представлены на рис. 7.2. Перед нами классический пример конкурентного исключения. Два близкородственных вида инфузорий Paramecium caudatum и Paramecium aurelia в раздельной культуре с постоянной средой и плотностью организмов, служащих нищей (инфузории питаются бактериями, которые в этих средах не размножаются, и поэтому бактерии вносили в среду через небольшие промежутки времени, поддерживая их плотность на постоянном уровне), росли в соответствии с типичной S-образной зависимостью, причем численность парамеций оставалась постоянной. Однако когда оба вида простейших выращивали в одной культуре, через 16 сут в ней обнаруживалась только Р. amelia. При этом ни один из организмов не нападал на другой и не выделял вредных веществ, просто популяции Р. aurelia отличаются более высокой скоростью роста (более высокой внутренней скоростью естественного роста), и поэтому при заданных условиях ограниченного количества пищи побеждают Р. caudatum (явная эксплуатационная конкуренция). Однако Paramecium caudatum и Paramecium bursaria могут выжить и достичь устойчивого равновесия при их совместном культивировании в одной и той же среде. Происходит это потому, что, хотя оба вида и питаются одной и той же пищей, Р. bursaria занимала часть культурального сосуда, где она могла питаться, не конкурируя с Р. caudatum. Таким образом, было показано, что местообитания у этих двух видов существенно различались, что позволяло им сосуществовать, несмотря на одинаковую пищу.
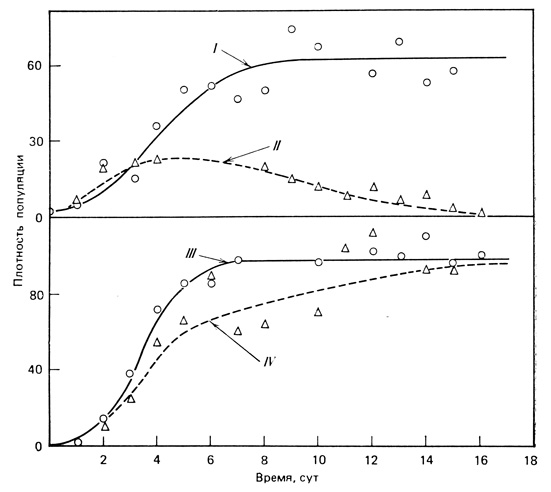
Рис. 7.2. Конкуренция между двумя близкородственными видами простейших, занимающих сходные ниши. В изолированных культурах при постоянном обеспечении пищей рост Paramecium caudatum (I) и Paramecium aurelia (III) происходит в соответствии с обычной S-образной зависимостью, но при совместном культивировании (II и IV) Р. caudatum (II) элиминируется. (По Гаузе из Allee et al., 1949)
Пример конкуренции с прямым подавлением описан Кромби (1947). Он обнаружил, что совместное культивирование в муке Tribolium и Oryzaephilus (другой род мучных хрущаков) заканчивается уничтожением Oryzaephilus, поскольку Tribolium активнее истребляет неполовозрелые стадии Oryzaephilus. Однако если в муку положить стеклянные трубочки, в которых могут прятаться неполовозрелые особи меньшего по размерам Oryzaephilus, то выживут обе популяции. Таким образом, если в среде имеются убежища, позволяющие скрываться от прямого воздействия (в данном случае хищничества), то конкуренция снижается настолько, что сохраняются оба вида.
Но довольно лабораторных примеров. Совершенно очевидно, что скученность в лабораторных экспериментах может быть весьма значительной, что приводит к чрезмерной конкуренции. В полевых исследованиях межвидовая конкуренция была подробно изучена у растений; результаты этих исследований позволили прийти к выводу (в настоящее время этот вывод является общепринятым), что конкуренция - важный фактор, вызывающий смену видов (как будет показано в гл. 8). Кивер (Keever, 1955) описал случай, когда залежь первого года была почти целиком занята чистым травостоем высокого сорняка, в дальнейшем его постепенно вытеснил отсюда другой вид, ранее не известный в этих местах. Эти два вида, хотя и принадлежат к разным родам, имеют очень сходные жизненные циклы (время цветения и созревания семян) и жизненные формы, попадали в условия интенсивной конкуренции. Тщательные последующие исследования этих залежных полей показали, что новый пришелец не вытеснял растущий здесь ранее вид; оказалось, что оба вида сосуществуют, но их численное соотношение зависит от почвы, времени и степени нарушений.
Мы уже отмечали, что конкуренция между особями одного вида - один из наиболее важных природных факторов, зависящих от плотности. То же самое можно сказать о межвидовой конкуренции. Конкуренция, по-видимому, оказывает огромное влияние на распределение близких видов, хотя зачастую об этом свидетельствуют лишь косвенные данные. Правило Гаузе трудно проверить в природе, используя стандартные методики, упрощающие реальную ситуацию, поскольку на исход конкуренции - сосуществование или исключение - могут влиять многие другие переменные. Тем не менее близкородственные виды или виды с очень сходными потребностями обитают обычно в разных географических областях или разных местообитаниях в одной и той же области либо избегают конкуренции каким-либо иным способом, например благодаря различиям в суточной или сезонной активности или различиям в пище. Вероятно, действие естественного отбора на протяжении длительного времени направлено на то, чтобы исключить или предотвратить продолжительную конфронтацию видов (гл. 8, разд. 4) со сходным образом жизни.
Эволюционный процесс, названный "смещением признаков", может быть причиной морфологических различий, усиливающих экологическое разобщение. Например, в Центральной Европе сосуществует шесть видов синиц (мелкие птицы рода Parus); их изоляция друг от друга обусловлена отчасти различиями в местообитаниях, отчасти же различиями в местах кормежки и размерах добычи, что находит отражение в небольших изменениях длины и толщины клюва. В Северной Америке встретить в одной местности более двух видов можно редко, хотя в целом на континенте обитает семь видов. Лэк (Lack, 1969) высказал предположение, что "американские виды синиц находятся на более ранних стадиях эволюции, чем европейские, и имеющиеся у них различия в строении клюва, размерах тела и пищевом поведении - это адаптации к соответствующим местообитаниям, а не адаптации, которые позволили бы им сосуществовать в одном местообитании".
Роль конкуренции в разделении местообитаний схематически иллюстрируется на рис. 7.3. Кривыми показаны границы местообитаний, внутри которых вид может существовать; отмечены также оптимальные и краевые зоны. При наличии конкуренции с близкородственными или экологически сходными видами диапазон местообитаний, возможных для данного вида, сокращается до оптимальных границ (т. е. вид распространяется в наиболее благоприятных для него местообитаниях, в которых он обладает какими-либо преимуществами по сравнению со своими конкурентами). Если же межвидовая конкуренция выражена слабо, то под влиянием внутривидовой конкуренции популяции данного вида расширяют границы своего местообитания.
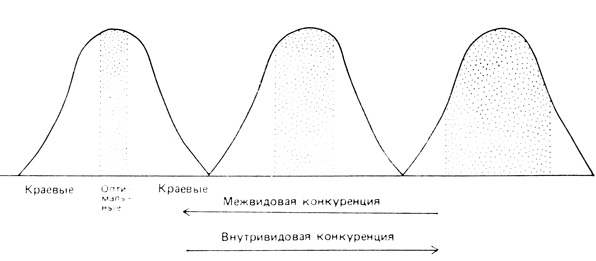
Рис. 7.3. Влияние конкуренции на распространение птиц в местообитаниях. Если преобладает внутривидовая конкуренция, то вид широко распространяется, занимая и менее благоприятные (краевые) области. Если же интенсивна межвидовая конкуренция, то распространение ограничивается узкими пределами, где имеются оптимальные условия. (Из Svardson, 1949, с изменениями)
На островах, которые не заселены потенциальными конкурентами, особенно хорошо прослеживается тенденция к расширению местообитаний. Например, на островах, где отсутствуют лесные полевки (Clethrionomys), луговые серые полевки (Microtus) часто занимают лесные местообитания (см. Cameron, 1964). Кроуэлл (Crowell, 1962) обнаружил, что на Бермудских островах кардиналы более многочисленны и занимают более краевые по условиям местообитания, так как здесь нет многих из его материковых конкурентов.
Хорошим примером экологического разделения сфер питания могут служить две родственные рыбоядные птицы Великобритании, большой баклан (Phalacrocorax carbo) и длинноносый, или хохлатый, баклан (Р. aristotelis), исследованные Лэком (Lack, 1945). Эти два вида обычно кормятся в одних и тех же водах и гнездятся на одних и тех же обрывах, однако более глубокий анализ показывает, что на самом деле места их гнездовий различаются, а, кроме того, существенные различия выявляются и в составе их пищи (рис. 7.4): длинноносый баклан ловит рыбу, плавающую в верхних слоях воды, тогда как большой баклан добывает пищу в основном у дна, где он ловит камбал и донных беспозвоночных (креветок и др.).
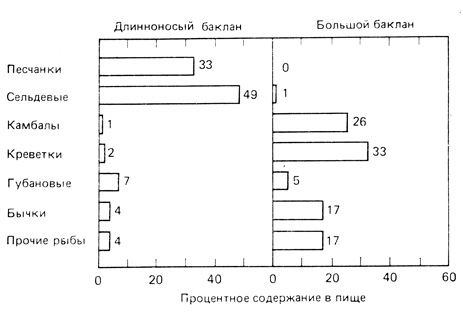
Рис. 7.4. Состав пищи двух близкородственных видов морских птиц, большого баклана (Phalacrocorax carbo) и длинноносого, или хохлатого, баклана (Р. aristotelis), живущих в период размножения в одних и тех же местах. Данные исследований показывают, что, несмотря на сходство местообитаний, пища этих бакланов различна. Таким образом, два вида занимают разные ниши и фактически не конкурируют за пищевые ресурсы. (Из Lack, 1945)
Тот факт, что близкородственные виды в природе четко разграничены, вовсе не означает, что для поддержания этой разобщенности между ними непрерывно должна поддерживаться конкуренция. В процессе эволюции у этих видов могут выработаться различные потребности и предпочтения, и тогда конкурентные отношения между ними прекращаются.
В Европе, например, один из видов рододендрона Rhododendron hirsutum растет на известковых почвах, а другой - R. farrugineum - на кислых; ни один из них не может жить на почве противоположного типа, поэтому конкуренция между ними практически отсутствует (Braun-Blanquet, 1932). Тил (Teal, 1958) экспериментально изучил выбор местообитания у манящих крабов (Uca), которые встречаются на засоленных маршах обычно порознь. Один вид U. pugilator был обнаружен на открытых песчаных отмелях, другой, U. pugnax, на илистом грунте, поросшем болотной травой. Тил выявил, что ни один из видов не стремится вторгнуться в местообитание другого, даже если оно и не заселено, потому что каждый из них может рыть норы только в предпочитаемом им субстрате. Отсутствие активной конкуренции, конечно, не означает, что ее следует исключить и в прошлом, когда она, по-видимому, служила фактором, который привел к выработке поведения, поддерживающего сейчас видовую изоляцию.
Три пробные модели, предложенные Филипом (Philip, 1955), могут служить основой для будущих наблюдений, анализа и экспериментальных работ: 1) несовершенная конкуренция, при которой межвидовая конкуренция служит лимитирующим фактором, но не приводит к полной элиминации одного из конкурентов из арены взаимодействия; 2) совершенная конкуренция, описываемая моделями Гаузе и Лотки - Вольтерра, при которой один вид постепенно элиминируется в процессе конкуренции за общий ресурс, и 3) сверхсовершенная конкуренция, при которой эффект подавления очень силен и проявляется немедленно, как, например, при выделении антибиотиков. Наглядный пример прямого подавления или "сверхсильной" конкуренции у растений приведен в следующем разделе.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'