Объяснения
О межвидовой конкуренции написано много и экологами, и генетиками, и эволюционистами. В большинстве случаев слово "конкуренция" применяется в ситуациях, в которых отрицательные взаимодействия являются следствием уменьшения ресурсов, используемых обоими видами. Более непосредственные взаимодействия, такие, как взаимное хищничество или выделение вредных веществ, следует отнести к другой категории (табл. 7.1), хотя для ее обозначения и нет общепринятого термина. Однако термин аллелопатия (введенный в разд. 4, гл. 2) теперь широко применяется для обозначения такого типа взаимодействия, когда один из конкурирующих видов выделяет химические посредники (Уиттакер (Whittaker, 1970) назвал их аллелопатическими веществами), которые обеспечивают конкурентное преимущество одного вида перед другим.
Конкурентное взаимодействие может касаться пространства, пищи или биогенных элементов, света, неиспользованных веществ, зависимости от хищников, подверженности болезням и т. д. и многих других типов взаимодействий. Результаты конкуренции представляют огромный интерес; они были многократно исследованы как один из механизмов естественного отбора. Межвидовая конкуренция, независимо от того, что лежит в ее основе, может привести либо к установлению равновесия между двумя видами, либо, при более жестокой конкуренции, к замене популяции одного вида популяцией другого, либо к тому, что один вид вытеснит другой в иное место или же заставит его перейти на использование иной пищи. Неоднократно отмечалось, что близкородственные организмы, ведущие сходный образ жизни и обладающие сходной морфологией, не обитают в одних и тех же местах. Если же они живут в одном месте, то часто используют разные ресурсы или активны в разное время. Объяснение экологического разделения близкородственных (или сходных в иных отношениях) видов получило известность как принцип Гаузе, по имени русского биолога, который в 1932 г. впервые подтвердил его существование экспериментально (см. рис. 7.2), или как принцип конкурентного исключения (Harden, 1940).
К числу наиболее широко обсуждавшихся в литературе теоретических проблем, относящихся к явлению конкуренции, относятся закономерности, получившие название уравнений Лотки - Вольтерра (эти модельные уравнения были предложены Лоткой и Вольтерра независимо друг от друга в 1925 и 1926 гг.). Эти уравнения представляют пару дифференциальных уравнений, сходных с уравнением, приведенным в предыдущем разделе. Такие уравнения можно использовать для моделирования систем хищник - жертва, паразит - хозяин, конкуренции и других типов взаимодействия между двумя видами. В случае конкуренции в ограниченном пространстве, где для каждой популяции имеется определенный уровень равновесия, характеризуемый величиной К, уравнения одновременного роста двух популяций, основанные на логистическом уравнении, можно записать так:
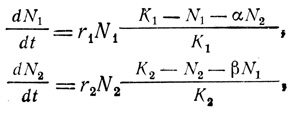
где N1 и N2 - соответственно численности видов 1 и 2. α - коэффициент конкуренции, характеризующий ингибирующее воздействие вида 2 на вид 1, β соответствует коэффициенту конкуренции, означающему ингибирование вида 2 видом 1. В более общем виде уравнения конкуренции были предложены Вигертом (Wiegert, 1974). В отсутствие убежищ (рефугий) или в других условиях вид, который сильнее ингибирует своего партнера, вытеснит его независимо от того, что коэффициенты конкуренции очень :малы по сравнению с соотношениями насыщающих плотностей (K1/K2 или K2/K1).
Чтобы понять причины конкуренции, необходимо рассматривать не только особенности популяций и условия, определяющие конкурентное исключение, но также ситуации, в которых сосуществуют сходные виды, поскольку в открытых природных системах большое число видов фактически пользуется общими ресурсами. В табл. 7.2 и на рис. 7.1 показана ситуация, которую можно было бы назвать моделью Tribolium-Trifolium; эта модель наглядно демонстрирует конкурентное исключение в паре видов жуков (Tribolium) и сосуществование двух видов клевера (Trifolium).
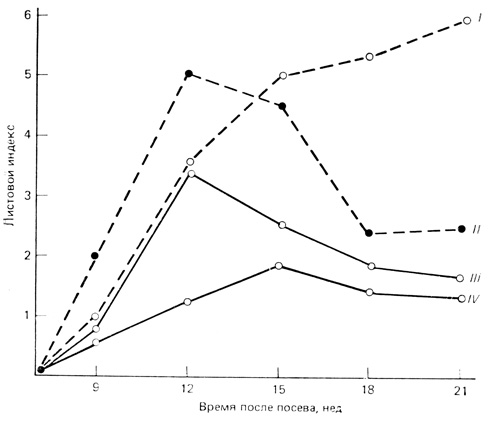
Рис. 7.1. Сосуществование видов клевера (Trifolium). На графике показан рост популяций двух видов в чистых (т. е. представленных только одним видом) и смешанных травостоях. Обратите внимание на то, что кривые роста двух видов в чистых травостоях различны и в разное время достигают зрелости. Это и другие различия позволяют видам сосуществовать в смешанных травостоях, но при этом их плотность понижена, так как они в одинаковой степени мешают друг другу. Листовой индекс, который используется для оценки плотности биомассы, представляет собой отношение площади поверхности листьев к поверхности почвы (см2/см2). I - Т. fragiferum, чистый травостой; II - Т. repens, чистый травостой; III - Т. repens, смешанный травостой; IV - Т. fragiferum, смешанный травостой

Таблица 7.2. Случай конкурентного исключения в популяциях мучного хрущака (Tribolium). (По Park, 1954)
1 (Каждый из 6 вариантов опытов проведен в 20-30 повторностях. В чистой культуре каждый вид выживает при любых сочетаниях условий, однако при совместном культивировании двух видов выживает только один. В процентах выражено относительное число повторностей, в которых сохранялся только один вид, а другой элиминировался.)
Одно из наиболее тщательных и длительных экспериментальных исследований межвидовой конкуренции было проведено в лаборатории покойного доктора Томаса Парка в Чикагском университете. Парк, его студенты и сотрудники работали с мучным хрущаком, в частности с видами рода Tribolium. Эти мелкие жуки могут завершить весь свой жизненный цикл в очень простой и однородной среде - в банке с мукой или пшеничными отрубями. В этом случае среда служит одновременно и пищей, и местообитанием личинок и имаго. Если регулярно добавлять свежую среду, то популяция жуков может поддерживаться долгое время. С точки зрения представлений о потоке энергии такую экспериментальную систему можно описать как стабилизированную гетеротрофную экосистему, в которой импорт энергии пищи уравновешивается затратами на дыхание.
Оказалось, что если в этот однородный мирок поместить два разных вида Tribolium, то раньше или позже один из видов неизбежно будет элиминирован, тогда как другой будет успешно размножаться. Один вид всегда "побеждает", или, иными словами, два вида Tribolium не могут выжить в этом специфическом "одноместном" микрокосме. Относительное число особей каждого вида, первоначально внесенных в культуру (т. е. исходное соотношение особей) не влияет на окончательный результат, однако "климат" экосистемы существенно сказывается на исходе борьбы. Как показано в табл. 7.2, один из видов (Т. castaneum) всегда побеждает в условиях высокой температуры и влажности, тогда как другой (Т. confusum) всегда побеждает в прохладном и сухом "климате", хотя и тот и другой при любом из испытанных сочетаний условий может неопределенно долго существовать в чистой культуре. В промежуточных условиях для каждого вида имеется определенная вероятность оказаться победителем (например, для Т. castaneum эта вероятность в теплой и влажной среде составляет 0,86). Понять некоторые результаты конкурентного взаимодействия помогает анализ свойств популяции каждого вида в чистой культуре. Например, было обнаружено, что победителем обычно оказывается вид, обладающий в изучаемых условиях намного большей скоростью роста (r), чем другой. Если же величины r для обоих видов различаются не слишком сильно, то вид с более высоким показателем не всегда окажется победителем. Вирусное заболевание в одной из популяций легко может решить исход дела. Финер (Feener, 1981) описал случай, когда конкурентное равновесие между двумя видами муравьев было нарушено паразитическим двукрылым. Кроме того, конкурентоспособность может в значительной степени различаться у различных генетических линий.
Пользуясь данными, полученными в модельных экспериментах на Tribolium, легко создать условия, в которых виды не исключали бы друг друга, а сосуществовали. Если культуры попеременно помещать то в жаркие и влажные, то в сухие и холодные условия (для имитации сезонных изменений погоды), преимущество одного вида перед другим будет недостаточно длительным для уничтожения другого. Если бы система культивирования была "открытой" и особей доминирующего вида вынуждали эмигрировать (или их бы удаляли, как это делает хищник) достаточно быстро, то конкуренция оказалась бы настолько слабой, что оба вида могли бы сосуществовать. Существованию могли бы благоприятствовать и многие другие условия.
Интересные эксперименты по конкуренции у растений провел Дж. Л. Харпер и его коллеги из Университетского колледжа Северного Уэльса (см. Harper, 1961; Harper и Clatworthy, 1963; Clatworthy, Harper, 1962). Результаты одного из этих исследований представлены на рис. 7.1. Вследствие различий в характере роста два вида клевера могут сосуществовать в одной и той же среде (при одинаковых условиях освещенности, температуры, на одной и той же почве и т. д.). Из этих двух видов Trifolium repens растет быстрее и скорее достигает максимальной облиственности. Однако Т. fragijerum имеет более длинные черешки и выше посаженные листья, поэтому он может перейти в верхний ярус раньше, чем быстро растущий вид (особенно после того, как скорость роста Т. repens пошла на убыль), и таким образом избежать затенения. В смешанных травостоях из-за этих особенностей каждый вид ингибирует развитие другого, но оба они оказываются в состоянии завершить жизненный цикл и дать семена, хотя плотность каждого вида понижена (впрочем, общая плотность в смешанных травостоях двух видов была примерно равна плотности в чистых травостоях). В этом случае оба вида, несмотря на сильную конкуренцию за свет, могут сосуществовать, и это сосуществование обусловлено особенностями морфологии и различиями во времени максимального роста. Харпер (Harper, 1961) пришел к заключению, что два вида растений могут длительно сосуществовать, если их популяции независимо регулируются посредством одного или нескольких из следующих механизмов: 1) различия потребностей в питании (например, бобовые и небобовые); 2) различия причин смертности (например, разная чувствительность к выеданию скотом); 3) чувствительности к различным токсинам и 4) чувствительности к одному и тому же регулирующему фактору (свет, вода и т. д.) в разное время (как в случае только что описанного примера с клевером).
Парк (Park, 1954), Брайен (Brian, 1956) и Кромби (Crombiey 1947) (все работали с мучными хрущаками) впервые стали проводить различие между непрямой конкуренцией, или эксплуатационной конкуренцией, и прямой конкуренцией, или интерференционной конкуренцией. На основании данных, полученных при изучении головастиков, Стейнуошер (Steinwasher, 1978) пришел к заключению, что эксплуатационная конкуренция сменяется интерференционной конкуренцией при уменьшении количества пищи. Хатчинсон (Hutchinson, 1978) полагает, что интерференционная конкуренция встречается все чаще по мере продвижения вверх по филогенетическому "древу" от примитивных фильтрующих простейших и ветвистоусых, которые обычно конкурируют за захват пищевых частиц, к позвоночным с их развитыми поведенческими механизмами агрессии, территориальности и т. д. Он заключает (как это сделал Слободкин (Slobodkin, 1964) на основе экспериментов по конкуренции у Hydra), что оба указанных типа конкуренции перекрываются, но что по теоретическим соображениям полезно различать эти два процесса, так как уравнениям эксплуатационной конкуренции соответствуют графики с прямоугольными изоклинами, тогда как для интерференционной конкуренции характерны криволинейные изоклины. Таким образом, сосуществование более вероятно во втором случае. Более подробные сведения, касающиеся конкуренции, можно найти в книге Хатчинсона (Hutchinson, 1978, гл. 4). Хороший обзор экспериментальных и генетических исследований, выполненных на популяциях Drosophila, имеется у Айалы (Ayala, 1972) и Ричмонда и др. (Richmond et al., 1975).
При изучении литературы, посвященной конкуренции, складывается общее впечатление, что в системах, где отсутствуют или понижены иммиграция и эмиграция, происходит более жестокая конкуренция и более вероятно конкурентное исключение. К числу таких систем следует отнести лабораторные культуры, острова или другие природные ситуации с трудно преодолимыми преградами для входа и выхода. В обычных природных открытых системах вероятность сосуществования выше.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'