Примеры
Сильные бури, внезапные падения температуры и другие резкие изменения физических факторов - наиболее характерные примеры независимых от плотности воздействий. Изучая в течение трех лет моллюска Acmaea, живущего на камнях в литоральной зоне, Фрэнк (Frank, 1965) обнаружил, что большая часть изменений, происходящих в популяции, зависит от плотности; исключение составляют случаи резкого увеличения смертности после суровых морозных зим, когда поверхность камней разрушается и удаляется вместе с моллюсками независимо от их численности. Хороший случай "совершенной" зависимости от плотности уже был нами рассмотрен на примере роста дрожжей (рис. 6.12). Как отмечалось в обсуждении, линейная зависимость от плотности маловероятна в природных открытых системах. Проведя тщательное исследование васильковой орехотворки, Варли (Varley, 1947) показал, что влияние ее основного паразита, насекомого Eurytoma curta, зависит от плотности, поскольку в условиях, когда плотность популяции хозяина велика, паразит уничтожает большее (как в процентном, так и в абсолютном выражении) число особей хозяина (табл. 6.4). Однако в другом исследовании Варли и Эдвардс (Varley, Edwards, 1957) сообщили, что когда "площадь поиска" мала, как в случае паразитической осы Mormoniella, влияние паразита на своего двукрылого хозяина не обязательно зависит от плотности. Таким образом, важную роль могут играть различия в поведении. Холдинг (Rolling, 1965, 1966) предложил серию математических моделей, в которые ввел поведенческие признаки. Эти модели позволяют прогнозировать эффективность регулирующего влияния данного насекомого-паразита на насекомое-хозяина при различных уровнях плотности.

Таблица 6.4. Зависимое от плотности влияние паразитического перепончатокрылого на хозяина - личинку галловой мухи. (По Varley, 1947)
Нередко наблюдается явная корреляция между теорией и той средой, которую изучает теоретик. Так, экологи, работающие в местностях с резко неблагоприятными условиями (например, в аридных областях) или имеющие дело с мелкими организмами (такими, как насекомые или планктон, для которых характерны короткие жизненные циклы, высокий биотический потенциал, высокая интенсивность метаболизма на 1 г массы, а следовательно, и небольшая биомасса на единицу пространства в единицу времени), прежде всего обращают внимание на: 1) значение периода времени, когда скорость прироста (r) положительна; 2) значение независимых от плотности факторов, таких, как погода, в определении продолжительности благоприятных периодов; 3) второстепенную роль внутрипопуляционных факторов самоограничения; 4) общее отсутствие стабильности в динамике плотности любых видов, даже в стабильных экосистемах. Эти моменты подчеркнуты в хорошо известной книге Андреварты и Бирча (Andrewartha, Birch, 1954).
Экологи, работающие в мягких, благоприятных условиях (таких, как английские сады, коралловые рифы или тропические леса) или с более крупными организмами (птицы, млекопитающие или лесные деревья, у которых жизненные циклы более продолжительны, а численность и биомасса более четко соответствуют потоку энергии), обращают внимание на: 1) значение зависимых от плотности факторов, и прежде всего - лимитирующее значение внутривидовой конкуренции (как в уравнении S-образного роста) и различных межвидовых регулирующих влияний (межвидовая конкуренция, паразиты и т. д.); 2) стабильность или по меньшей мере постоянство характера изменений плотности; 3) значение биологических механизмов регуляции. Характерно, что исследователи, работающие с отдельными замкнутыми популяциями мелких организмов, такими, как культуры бактерий или лабораторные популяции мучных хрущаков, также обращают особое внимание на эти последние аспекты, что, в общем, неудивительно, поскольку биомасса на единицу объема в культурах больше, чем это обычно наблюдается в природе. В равной мере не следует удивляться и тому, что на исследователей, работающих с монокультурами зерновых и древесных растений, не производит впечатления эффективность биологического контроля.
Анализ теорий зависимой от плотности регуляции содержится в основополагающих работах Никольсона (Nicholson, 1954, 1957, 1958), Лэка (Lack, 1954); см. также приложение в работе Lack, 1966.
Все аспекты проблемы регулирования популяций можно представить в виде графической модели, приведенной на рис. 6.18. В основу графика положены данные о динамике популяции конкретного вида (австралийской листоблошки сем. Psyllidae, живущей на эвкалиптах), однако он иллюстрирует многие принципы, обсуждавшиеся выше. В норме наблюдается стабилизация популяций на уровне, соответствующем низкой численности (уровень 7, время I на рис. 6.18); этот уровень намного ниже уровня 2, которого достигает популяция при использовании всех пищевых и пространственных ресурсов. Регуляция на уровне ниже насыщающего осуществляется метеорологическими условиями и независимыми от плотности паразитизмом и хищничеством на, нимфах этого насекомого, а также зависимым от плотности хищничеством птиц на взрослых особях. Иногда стабилизирующие процессы ослабевают; обычно это происходит под влиянием необычайно низких температур, вызывающих снижение относительной численности паразитирующих организмов, в результате чего плотность увеличивается выше контролируемого уровня (т. е. "ускользает" от контроля; время II на рис. 6.18). Тогда происходит стремительный рост (время III) по J-образной кривой вследствие того, что 1) паразитизм на нимфах становится неэффективным из-за быстрого увеличения сверхпаразитов (паразитов, которые паразитируют на паразитах листоблошки) и 2) численность насекомоядных птиц растет медленнее, чем численность насекомых (вспомните значительные различия рождаемости у насекомых и птиц). Неограниченный какое-то время рост прекращается на уровне 2, когда нимфы выедают всю пищу, а взрослые особи не находят более мест, пригодных для откладки яиц. Затем, когда деревья остаются без листьев, а популяции хищников (птиц и муравьев) и паразитов возрастают, происходит резкий спад численности листоблошек (время IV). Если их численность опускается ниже уровня 1, то популяция опять попадает в зону действия механизмов регуляции, и ее численность в течение ряда лет, вероятно, будет небольшой (рис. 6.18, а). Если же плотность не снизится ниже контролируемого уровня, то может возникнуть новая внезапная вспышка численности (рис. 6.18, б).
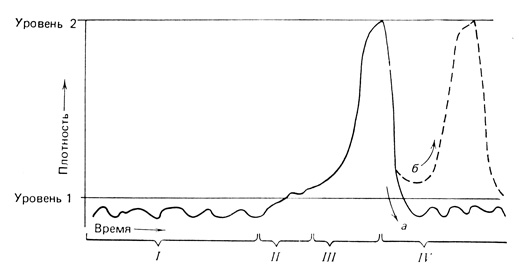
Рис. 6.18. Динамика численности насекомого Cardiaspina albitextura (Psyllidae), которое кормится на деревьях Eucalyptus blakelyi. В норме плотность популяции при совместном воздействии зависимых и независимых от плотности факторов (погоды, паразитов и хищников) стабилизируется на низком уровне. Иногда популяция ускользает от естественного контроля и вырывается на очень высокий уровень плотности; следствием этого является дефолиация деревьев на обширных участках. При плотности ниже уровня 1 эффективно действует механизм природного контроля. При плотности на уровне 2 строго лимитирующими становятся пища и места для откладки яиц. (Из Clark, 1964, см. также Clark et al., 1967, с. 158)
Эта модель иллюстрирует также некоторые трудности, которые возникают в практике контроля вспышек численности насекомых-вредителей. Частота вспышек увеличивается в результате хозяйственной деятельности человека, который, подобно "необычным" погодным условиям, нарушает работу природных регулирующих механизмов. Нередко численность популяции возрастает так быстро, что это обнаруживается лишь в фазе экспоненциального роста, когда проводить обработку уже слишком поздно. Очевидно, контролирующие мероприятия могут предотвратить вторую вспышку, если обработка была направлена на уничтожение именно данного насекомого-вредителя. Однако применение инсектицидов с широким спектром действия в период IV может принести больше вреда, чем пользы, поскольку при этом будут уничтожены также паразиты и хищники, что скорее увеличит, а не уменьшит вероятность новой вспышки. Во многих случаях, если нам не известны фаза популяционного цикла или состояние других популяций, участвующих в функционировании механизма естественного контроля, лучше воздержаться от каких бы то ни было истребительных мероприятий.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'