Объяснения
Популяция, как это хорошо выразил один из пионеров современной популяционной экологии Томас Парк (см. Allee et al., 1949), обладает "биологическими свойствами", присущими как популяции, так и составляющим ее организмам, и "групповыми свойствами", присущими только группе в целом. Биологические свойства характеризуют жизненный цикл популяции: популяция, так же как и отдельный организм, растет, дифференцируется и поддерживает сама себя. Популяция имеет определенную организацию и структуру, которые можно описать. В отличие от этого групповые свойства, такие, как рождаемость, смертность, возрастная структура и генетическая приспособленность, могут характеризовать только популяцию в целом. Таким образом, особь рождается, стареет, умирает, но применительно к особи нельзя говорить о рождаемости, смертности, возрастной структуре - характеристиках, имеющих смысл только на групповом уровне.
Ниже даются определения и краткие описания основных свойств популяции.
Плотность. Плотность популяции - это величина популяции, отнесенная к некоторой единице пространства. Ее обычно измеряют и выражают числом особей или биомассой популяции на единицу площади или объема, например 500 деревьев на 1 га, 5 млн. диатомей на 1 м3 воды или 200 кг рыбы на 1 га поверхности водоема. Иногда бывает важно различать среднюю плотность, т. е. численность (или биомассу) на единицу всего пространства, и удельную, или экологическую, плотность, т. е. численность (или биомассу) на единицу обитаемого пространства (доступной площади или объема, которые фактически могут быть заняты популяцией). Часто важнее знать не конкретную величину популяции в тот или иной момент времени, а ее динамику, т. е. возрастает она или уменьшается. В таких случаях полезны показатели относительного обилия. Они могут быть отнесены к определенным: промежуткам времени, например число птиц, отмеченных в течение часа. Их можно выразить также в процентах, например, частота встречаемости - это процент проб, в которых обнаружен данный вид. В описательных работах по геоботанике плотность и частоту встречаемости часто объединяют, получая показатель значимости для каждого вида.
На рис. 6.1 и в табл. 6.1 показано соотношение между плотностями популяций млекопитающих, их трофическими (энергетическими) уровнями и величиной особей. Хотя плотности популяций разных представителей млекопитающих как класса охватывают диапазон почти в пять порядков величины, диапазон колебаний плотности популяции для каждого вида или трофической группы намного меньше (рис. 6.1). Чем ниже трофический уровень, тем выше плотность, а чем крупнее животные внутри данного уровня, тем больше их биомасса. Так как у крупных животных интенсивность метаболизма на единицу массы меньше, чем у мелких, на данной энергетической базе может поддерживаться большая биомасса крупных животных.

Рис. 6.1. Диапазон изменений плотностей популяций (выраженных в биомассе на гектар) у разных видов млекопитающих. Результаты получены в предпочитаемых данным видом местообитаниях, где антропогенное влияние выражено незначительно. Виды расположены в соответствии с их трофическим уровнем, а в пределах каждого уровня - в соответствии с размерами животного, что иллюстрирует влияние на ожидаемую биомассу размеров организма и его положения в пищевой цепи. (Схема составлена по данным Mohr, 1940, и дополнена результатами позднейших исследований)
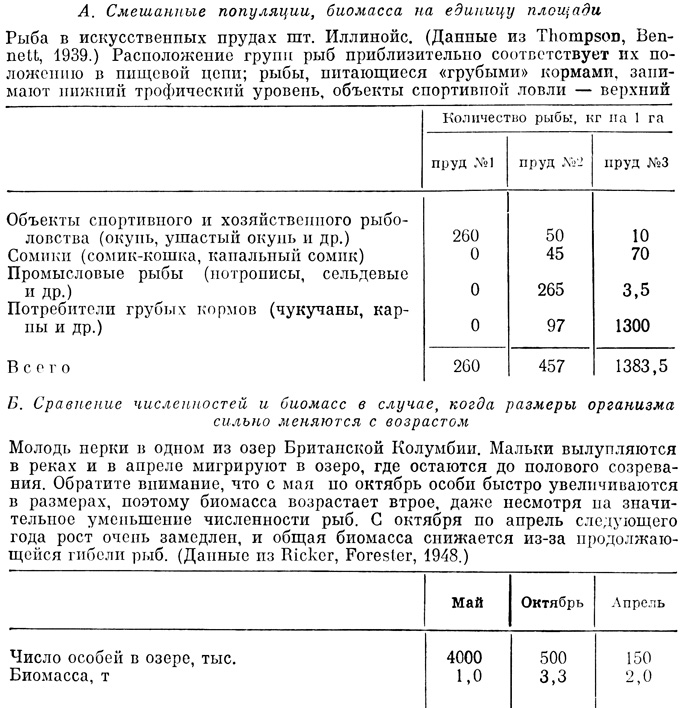
Таблица 6.1. Влияние трофического уровня на биомассу рыб (А) и взаимосвязь между численностью и биомассой в популяции с быстро меняющейся возрастной и размерной структурой (Б)
Если размеры особей в популяции и интенсивности их метаболизма примерно одинаковы, то плотность удобно выражать числом особей, однако часто встречается ситуация, показанная в табл. 6.1, Б. Относительные достоинства таких показателей, как численность, биомасса и поток энергии, обсуждались в гл. 3 (см., в частности, табл. 3.6). Вспомним следующее утверждение из этой главы: "данные по численности приводят к переоценке значения мелких организмов, а данные по биомассе - к переоценке роли крупных организмов". Поток энергии "служит более подходящим показателем для сравнения любого компонента с другим и всех компонентов экосистемы между собой".
Многие специальные количественные показатели и термины широко применяются только при работе с определенными популяциями или группами популяций. Лесные экологи, например, часто пользуются термином "площадь оснований" (суммарная площадь поперечных срезов стволов) как мерой плотности деревьев. Лесники же определяют число бордсовых, или досковых, футов на 1 акр как меру полезной с коммерческой точки зрения части всего древостоя. Эти и другие меры могут служить оценками плотности, поскольку они в известной степени отражают размеры "урожая на корню" и, следовательно, соответствуют данному ранее определению плотности в широком смысле.
Мы уже указывали, что если необходимо знать характер изменения популяции или если в данных условиях нельзя определить абсолютную плотность, то часто бывают полезными оценки относительного обилия. Термины "обильный", "обычный", "редкий" и т. п. применимы в тех случаях, когда они соответствуют измерениям или оценкам, полученным таким способом, что сравнение имеет смысл.
Легко догадаться, что "индексы" относительного обилия широко используются для популяций крупных животных и наземных растений, когда необходимо проводить оценку численности на больших площадях без чрезмерных затрат времени и денег. Например, административные органы, на которые возложена обязанность ежегодно регулировать охоту на перелетную водоплавающую дичь, чтобы соблюсти интересы и охотников, и птиц, должны знать, в каких местах популяции стали меньше, больше или остались такими же, как в предыдущем году. Это можно сделать путем оценки относительного обилия, полученной в результате обследования угодий егерями, опроса и учета гнездовий. Данные суммируют и представляют как число птиц, увиденных или убитых за единицу учетного усилия. Показатели, выраженные в процентах, широко используются при изучении растительности; общеупотребительны такие специальные показатели, как частота - процент пробных площадок, на которых встречается данный вид; обилие - процент особей в пробе; покрытие - процент поверхности почвы, оказавшейся закрытой при проекции на нее надземных частей растений. Следует быть осторожным и не путать эти индексы с истинной плотностью, которая всегда выражается по отношению к определенному пространству.
Различие между средней плотностью и экологической плотностью можно иллюстрировать данными Коля (Kahl, 1964) по лесному аисту в Национальном парке Эверглейдс (Флорида). На рис. 6.2 показано, что в этой местности плотность мелких рыб с падением уровня воды в течение сухого зимнего сезона в целом снижается, но экологическая плотность при этом возрастает, так как по мере сокращения зеркала воды увеличивается число рыб, приходящихся на единицу водной поверхности. Аисты откладывают яйца в такое время, что вылупление птенцов приходится на период пика экологической плотности рыб. Это облегчает родителям ловлю рыбы, которая составляет основную пищу птенцов. В мире лесного аиста в расчет принимается экологическая, а не средняя плотность пищевых объектов. В мире людей наблюдается во многом сходная картина: в расчет принимаются главным образом высококачественная энергия и пищевая продукция, сконцентрированные на 15 или 20 процентах земли, где возможен высокий урожай, но не рассеянные и трудно доступные для добывания ресурсы.
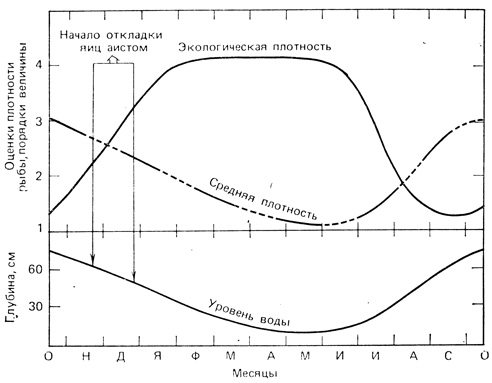
Рис. 6.2. Экологическая и средняя плотность рыб, служащих добычей аисту в период его размножения. По мере падения уровня воды в течение сухого сезона в водоемах на юге Флориды средняя плотность мелких рыб уменьшается (т. е. уменьшается число рыб на 1 км2 поверхности), так как сокращаются размеры и число водоемов, но экологическая плотность (т. е. количество рыбы на 1 м2 сохранившейся водной поверхности) возрастает, поскольку рыба скапливается в водоемах с меньшей площадью. Гнездование аиста приурочено к тому времени, когда максимальная доступность пищи совпадает с наибольшими пищевыми потребностями подрастающих птенцов. (По Kahl, 1964)
Для измерения плотности популяций испытано много разных методов, и сама по себе методология составляет важную область исследований. Было бы нецелесообразно подробно рассматривать здесь эти методы; их лучше постигать, пользуясь методическими и полевыми руководствами, а еще лучше советоваться с опытным исследователем, который вначале изучил оригинальную литературу, а потом модифицировал и усовершенствовал существующие методы, приспособив их к конкретной ситуации. Ничто не может заменить опыт, приходящий при проведении полевых исследований.
Практикуемые методы можно подразделить на несколько обширных категорий:
1. Тотальный учет, возможный иногда для крупных, хорошо заметных организмов (например, бизоны на открытых равнинах) или для организмов, собирающихся на период размножения в большие группы (например, морские птицы и тюлени).
2. Метод пробных площадок. Этот метод состоит в подсчете и взвешивании организмов на пробных площадках или на трансектах. Размеры и число пробных площадок должны быть достаточными для получения оценки численности в обследуемой территории.
3. Методы мечения с повторным отловом (для подвижных животных). Из популяции отлавливают часть животных, метят их и выпускают. Доля меченых животных в последующей выборке используется для определения общей численности (см. Zippin, 1958).
4. Метод изъятия, при котором число организмов, собранных с некоторой площади при последовательных выборках, откладывается по оси ординат, а число из собранных ранее - по оси абсцисс. Если вероятность поимки относительно постоянна, то точки ложатся вдоль прямой линии. Эту линию можно продлить до нулевой точки (на оси абсцисс), теоретически соответствующей 100%-ному изъятию с данной площади (см. Menhinick, 1963).
5. Методы без взятия проб (применимые к неподвижным организмам, таким, как деревья). Примером может служить метод случайных точек, при котором измеряют расстояние от ряда произвольно выбранных точек до ближайших особей по всем четырем направлениям. Плотность на единицу площади оценивают по среднему расстоянию (см. Phillips, 1959).
Рождаемость. Рождаемость - это способность популяции к увеличению численности. Термин рождаемость используется также и при анализе популяций человека (в демографии)*. Фактически в популяционной экологии это просто более широкий термин; он характеризует появление на свет новых особей любого организма независимо от того, рождаются ли они, вылупляются из яиц, прорастают из семян или появляются в результате деления. Максимальная рождаемость (иногда ее называют абсолютной или физиологической) - это теоретический максимум скорости образования новых особей в идеальных условиях (когда отсутствуют лимитирующие экологические факторы, размножение сдерживается только физиологическими). Максимальная рождаемость постоянна для данной популяции. Экологическая, или реализованная, рождаемость (или просто "рождаемость" без какого-либо определения) обозначает увеличение численности популяции при фактических или специфических условиях среды. Эта величина не постоянна и варьирует в зависимости от размерного и возрастного состава популяции и физических условий среды. Обычно рождаемость выражают либо как скорость, определяемую путем деления общего числа вновь появившихся особей на время (абсолютная или общая рождаемость), либо как число вновь появившихся особей в единицу времени на 1 особь в популяции (удельная рождаемость).
* (В оригинале сказано, что для обозначения рождаемости экологи и демографы используют разные термины (соответственно "natality" и "birth rate"). В русской литературе в обоих случаях используется один термин. - Прим. перев.)
Различие между абсолютной и удельной рождаемостью легко проиллюстрировать на таком примере: предположим, что популяция из 50 простейших в некотором объеме воды увеличивается путем деления. Через час численность ее возросла до 150 особей. Абсолютная рождаемость при этом равна 100 особям в час, а удельная рождаемость (средняя скорость изменения численности на особь в популяции) составляет 2 особи в час при 50 исходных. Или же предположим, что в городе с населением 10000 появилось 400 новорожденных. Абсолютная рождаемость составит 400 в год, а удельная - 0,04 (4 на 100, или 4%). В демографии рождаемость принято рассчитывать на одну женщину репродуктивного возраста, а не на все население.
Различие между максимальной и реализованной рождаемостью можно проиллюстрировать данными, полученными Лески (Laskey, 1939) при изучении природной популяции певчей птицы Sialis sialis на искусственных гнездовьях городского парка в Нашвилле, Теннесси, и данными, полученными Парком (Park, 1934) на лабораторной популяции мучного хрущака. Птицы отложили 510 яиц (около 15 яиц на самку в трех последовательных кладках); это число соответствует максимальной рождаемости. Оперилось всего только 265 птенцов, поэтому экологическая или реализованная рождаемость составляет 52% максимальной (около 8 птенцов на самку или 4 на гнездящуюся пару). В опытах на мучном хрущаке были получены иные данные; так, в одном из контейнеров жуками было отложено 12 000 яиц (см. Park, Ginsburg, Horwitz, 1945), из которых вылупились только 773 (или 6%) личинки. В общем для видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая потенциальная и низкая реализованная рождаемость. В последующих разделах будут обсуждаться другие обстоятельства, влияющие на рождаемость.
Смертность. Смертность отражает гибель особей в популяции. В определенной степени это понятие является антитезой рождаемости. Смертность эквивалентна уровню смертности в демографии. Подобно рождаемости, смертность можно выразить числом особей, погибших за данный период (число смертей в единицу времени), или же в виде удельной смертности для всей популяции или любой ее части. Экологическая, или реализованная, смертность - гибель особей в данных условиях среды. Эта величина, как и экологическая рождаемость, не постоянна и изменяется в зависимости от условий среды и состояния самой популяции. Теоретическая минимальная смертность - величина, постоянная для популяции; она представляет собой гибель особей в идеальных условиях, при которых популяция не подвергается лимитирующим воздействиям. Даже в самых лучших условиях особи будут умирать от старости. Этот возраст определяется физиологической продолжительностью жизни, которая, конечно же, часто намного превышает среднюю экологическую продолжительность жизни. Нередко значительно больший интерес представляет не смертность, а выживаемость. Если число погибших особей выразить через М, то выживаемость будет равна 1-М.
Как и рождаемость, смертность, особенно у высших организмов, широко варьирует с возрастом. В связи с этим большое значение имеет определение удельной смертности для возможно большего числа возрастных групп или стадий развития, поскольку это позволяет экологам выяснить механизмы, определяющие общую смертность в популяции. Полная картина смертности в популяции последовательно описывается статистическими таблицами выживания, специально разработанными в демографических исследованиях. Раймонд Пирл впервые использовал таблицы выживания для решения некоторых проблем общей биологии, применив их к данным, полученным в лабораторных исследованиях плодовой мушки Drosophila (Pearl, Parker, 1921). Диви (Deevy, 1947, 1950) обобщил в виде таких таблиц данные по многим природным популяциям от коловраток до горного барана. Со времени появления обзоров Диви таблицы выживания были составлены для самых разных природных и экспериментальных популяций. В табл. 6.2 и 6.3 суммированы данные, полученные для аляскинской популяции дикого горного барана, популяции рисового долгоносика и одной гипотетической популяции. В табл. 6.3 включена также удельная рождаемость для разных возрастов. Возраст баранов определяли по рогам (чем старше баран, тем больше костяных колец). Когда барана убивает волк или он гибнет от других причин, его рога сохраняются еще долгое время. Адольф Мьюри в течение нескольких лет изучал взаимоотношения волка и дикого барана в национальном парке Мак-Кинли на Аляске. Он собрал много рогов, получив, таким образом, интересные данные о возрасте, в котором гибнут бараны в природных условиях, когда они подвержены опасностям, в том числе и нападению хищников - волков (человек их не трогает постольку, поскольку охота на барана в Национальном парке Мак-Кинли запрещена).
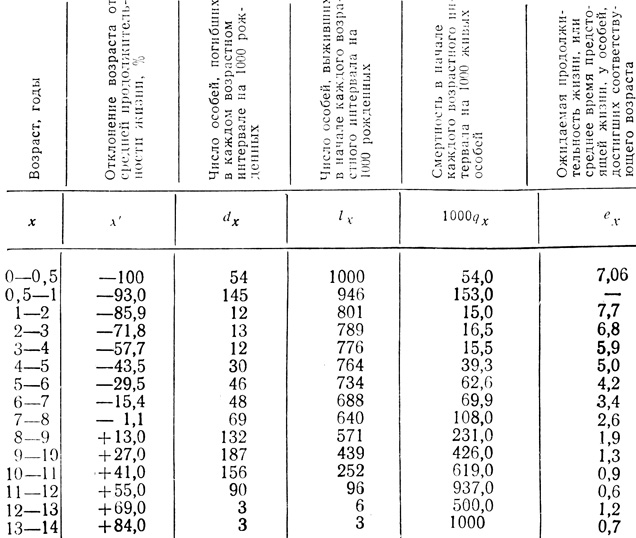
Таблица 6.2. Таблица выживания для популяции горного барана Далла (Ovis d. dalli) (Данные Murie, 1944, из Deevey, 1947), построенная на основе данных о возрасте 608 особей к моменту смерти. Учитывались бараны, погибшие до 1937 г. Данные для обоих полов объединены. Средняя продолжительность жизни - 7,09 лет. Небольшое число черепов было лишено рогов, но, по остеологическим данным, они принадлежали особям 9 лет и старше и поэтому были распределены пропорционально среди старших возрастных групп
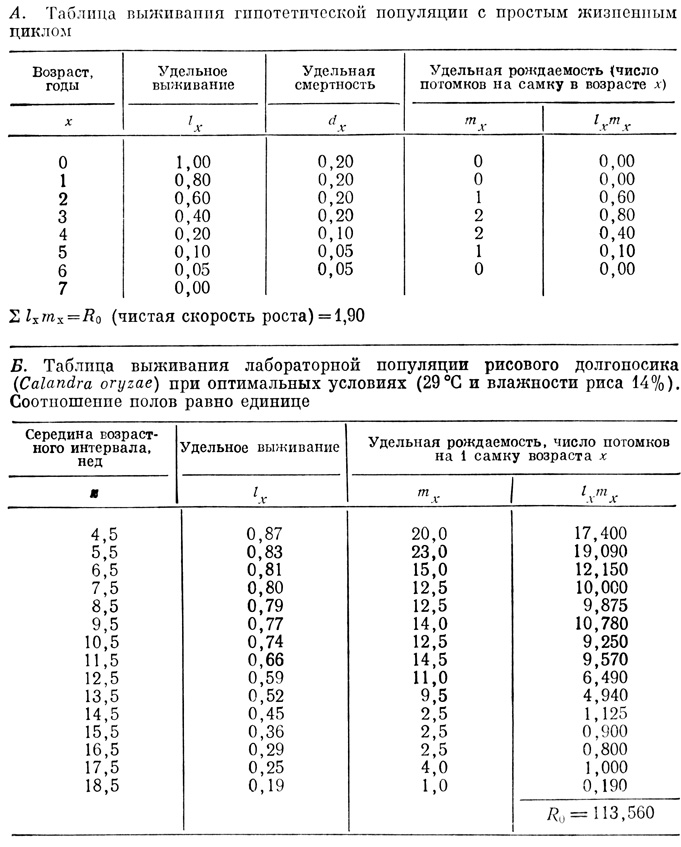
Таблица 6.3. Таблицы выживания с удельными величинами выживания и рождаемости
Таблица выживания состоит из нескольких столбцов, обозначенных следующим образом: lх - число особей из 1000 (или любого другого удобного числа) представителей данной популяции, которые выживают к концу определенного интервала времени (день, месяц, год и т. д.), указываемого в столбце х; dx - число особей, погибших в последовательные интервалы времени; qx - смертность в течение последовательных интервалов по отношению к величине исходной популяции и ех - ожидаемая продолжительность жизни в конце каждого интервала. Как показано в табл. 6.2, средний возраст горных баранов превышает 7 лет, и если баран смог выжить в течение первого года, то его шансы на выживание до относительно позднего возраста, несмотря на обилие волков и другие превратности судьбы, остаются довольно высокими.
Кривые, построенные на основе таблиц выживания, могут быть весьма информативными. Если данные столбца lx отложены на графике, где горизонтальная координата соответствует интервалу времени, а вертикальная - числу выживших, то полученная кривая называется кривой выживания. Такие кривые, построенные в полулогарифмическом масштабе, причем интервалы времени по оси абсцисс отложены как процент средней продолжительности жизни (см. столбец x' в табл. 6.2) или как процент общей продолжительности жизни, позволяют сравнивать виды с очень разной продолжительностью жизни. Кроме того, прямая линия, построенная в полулогарифмическом масштабе, свидетельствует о постоянстве удельного выживания.
Кривые выживания подразделяют на три общих типа, показанных на рис. 6.3. Сильно выпуклая кривая (I на рис. 6.3) характерна для таких видов, как, например, баран Далла, в популяциях которых смертность почти до конца жизненного цикла остается низкой. (Мы предлагаем нанести данные из столбца lx табл. 6.3 на полулогарифмическую миллиметровую бумагу и сравнить форму полученной кривой с кривыми на рис. 6.3.) Этот тип кривой выживания характерен для многих видов крупных животных и, конечно, для человека. Другой крайний вариант, сильно вогнутая кривая (V на рис. 6.3), получается, если смертность высока на ранних стадиях. Хорошей иллюстрацией этого служат устрицы и другие двустворчатые моллюски, а также дубы: смертность очень велика у свободноплавающих личинок и прорастающих желудей, но, как только особь хорошо приживется на подходящем субстрате, ожидаемая продолжительность жизни сильно увеличивается. К промежуточному типу относятся кривые выживания для тех видов, у которых удельная выживаемость для каждой возрастной группы более или менее одинакова, так что в полулогарифмическом масштабе кривые приближаются к диагоналям (III, IV на рис. 6.3). Ступенчатый тип кривой выживания характерен для видов, у которых выживаемость сильно варьирует на последовательных стадиях жизненного цикла, как это часто бывает у насекомых с полным превращением (голометаболических), например у бабочек. На изображенной кривой (II на рис. 6.3) крутые участки соответствуют стадиям яйца, окукливания и короткоживущему имаго; пологие участки соответствуют стадиям личинки и куколки, у которых смертность ниже (см. Itô, 1959). Вероятно, в природе не существует популяций, у которых выживаемость постоянна на протяжении всего жизненного цикла (кривая III на рис. 6.3); однако слегка вогнутые или сигмоидные кривые (IV на рис. 6.3) всегда характерны для многих птиц, мышей и кроликов. В этих случаях смертность высока у молодых особей, а у взрослых (в возрасте 1 года и старше) она ниже и более или менее постоянна.
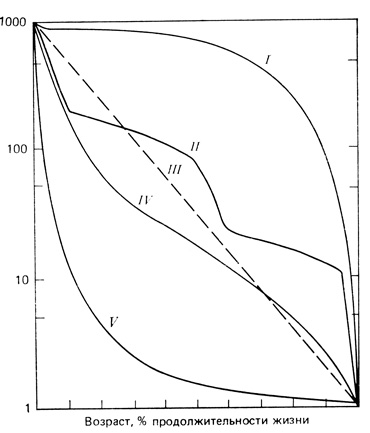
Рис. 6.3. Разные типы кривых выживания. По оси ординат отложено число выживших на тысячу особей (логарифмическая шкала), а по оси абсцисс - возраст в процентах от продолжительности жизни. I - выпуклая кривая (наибольшая смертность приходится на конец жизни). II - ступенчатая кривая (выживание резко меняется при переходе от одной стадии жизненного цикла к другой). III - теоретическая кривая (прямая; выживание для разных возрастов остается постоянным). IV - сигмоидная кривая, приближающаяся к III. V - вогнутая кривая (смертность очень высока на ранних стадиях)
Форма кривой выживания связана со степенью заботы о потомстве и другими способами защиты молоди. Так, кривые выживания пчел и дроздов (которые заботятся о потомстве) значительно менее вогнуты, чем у кузнечиков и сардин (которые не заботятся о потомстве). У последних видов это обстоятельство, без сомнения, компенсируется значительно большим числом откладываемых яиц (отношение максимальной к реализованной рождаемости велико, как отмечалось в предыдущем разделе).
Форма кривой выживания очень часто варьирует также и в зависимости от плотности популяции. На рис. 6.4 показаны кривые выживания двух популяций оленя, обитающих в зарослях чапараля в Калифорнии. Кривая выживания более плотной популяции заметно вогнута. Иными словами, у оленей, живущих на освоенной территории, где количество пищи больше в результате планомерного выжигания старых зарослей, ожидаемая продолжительность жизни меньше, чем у оленей, живущих на неосвоенной территории; это, по-видимому, связано с более интенсивной охотой, внутривидовой конкуренцией и т. д. С точки зрения охотника, освоенная территория более благоприятна, но с точки зрения оленя, на менее населенной территории больше шансов выжить. То же самое можно сказать и относительно человеческих популяций, где высокая плотность не всегда бывает благоприятна для отдельного индивидуума. Многие экологи считают, что быстрый рост и высокая плотность в популяциях человека опасны не столько для выживания, сколько для качества жизни индивидуума. И это несмотря на то, что "экологическая" продолжительность жизни человека значительно увеличилась благодаря достижениям современной медицины, улучшению питания и т. д. Кривая выживания для человека приближается к остроугольному типу с минимумом смертности. Однако максимальная, или "физиологическая", продолжительность жизни человека, по-видимому, не увеличилась, поскольку в наше время до 100 лет доживает не больше людей, чем в минувшие столетия. Эти парадоксы еще будут обсуждаться в гл. 8.
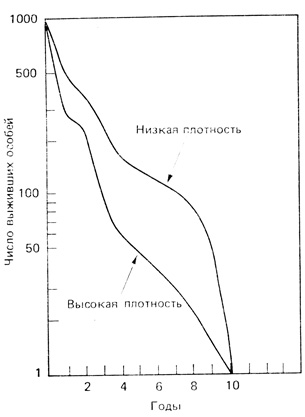
Рис. 6.4. Кривые выживания для двух стабильных популяций чернохвостого оленя, обитающих в зарослях чапараля в Калифорнии. Популяция с высокой плотностью (примерно 40 оленей на 1 км2) обитает на территории, где покров открытых кустарников и травянистых растений путем регулярных палов постоянно поддерживается в таком состоянии, что имеется большое количество новых побегов, которые олени обгрызают. Популяция с низкой плотностью (примерно 17 оленей на 1 км2) обитает на территории, где не ведется хозяйства; старые заросли кустарников не выжигались уже 10 лет. Свежие гари могут прокормить до 57 оленей на 1 км2, но эта популяция нестабильна, и поэтому кривые выживания нельзя строить по данным о возрастном распределении. (По Taber и Dasmann, 1957)
Чтобы подготовить основу для создания математических моделей роста популяций, которые будут рассматриваться в следующих разделах, целесообразно дополнить таблицы выживания данными по удельной рождаемости (числом потомков, производимых самкой в единицу времени) в каждом возрастном классе. В табл. 6.3 это столбец mx, так что теперь это не просто таблица "смертей".
Если перемножить величины lx и mx и результаты для разных возрастных классов сложить, то получится чистая скорость размножения (R0).
Таким образом,
(в данном случае lx относится только к самкам). Величина чистой скорости размножения 1,9 в табл. 6.3 означает, что число потомков на каждую самку составляет 1,9. Если число самок равно числу самцов, то получается, что примерно за одно поколение популяция полностью обновляется. Лабораторная популяция рисового долгоносика при самых оптимальных условиях увеличивается через каждое поколение в 113,6 раз (т. е. R0 = 113,6; табл. 6.3, Б). В природе в стабильных условиях величина R0 для популяции в целом должна составлять около 1. Перис и Пителка (Paris, Pitelka, 1962), использовав таблицы выживания и данные по lx и mx по годовым классам, высчитали величину R0 для популяции мокриц в злаковнике. Она оказалась равной 1,02, что свидетельствует о приблизительном равновесии между числом рождений и числом смертей.
Особенности размножения сильно влияют на рост популяции и другие ее характеристики. Естественный отбор может воздействовать на разные типы изменений жизненного цикла, что приводит к появлению адаптивных свойств. Так, давление отбора может изменить сроки начала размножения, не затрагивая общее число производимых потомков, или же отбор может влиять на продукцию, или "величину выводка", не меняя сроков размножения. Эти и многие другие аспекты динамики популяции можно выявить при анализе таблиц выживания.
Возрастной состав популяции. Возрастная структура популяции является ее важной характеристикой, которая, как было показано на примерах, обсуждавшихся в предыдущем разделе, влияет как на рождаемость, так и на смертность. Соотношение разных возрастных групп в популяции определяет ее способность к размножению в данный момент и показывает, чего можно ожидать в будущем. Обычно в быстро растущих популяциях значительную долю составляют молодые особи, в популяции, находящейся в стационарном состоянии, возрастное распределение более равномерно, а в популяции, численность которой снижается, будет содержаться большая доля старых особей (рис. 6.5 и 6.6). Возрастная структура популяции может изменяться и без изменения ее численности. По имеющимся данным, для каждой популяции характерно "нормальное", или стабильное, возрастное распределение, к которому направлены изменения реальных распределений; теоретически это впервые предсказал Лотка (Lotka, 1925). Коль скоро достигается стабильное возрастное распределение, временные отклонения от него могут быть вызваны лишь, необычными увеличениями рождаемости или смертности, после чего происходит спонтанный возврат к стабильной ситуации. По мере развития человеческого общества от состояния освоения новых территорий и быстрого роста численности к зрелому состоянию стабильной популяции процент населения в младших возрастных классах уменьшается (рис. 6.6). Это изменение возрастной структуры и увеличение процента населения старших возрастных классов оказывает глубокое воздействие на жизненный уклад общества и его экономику.
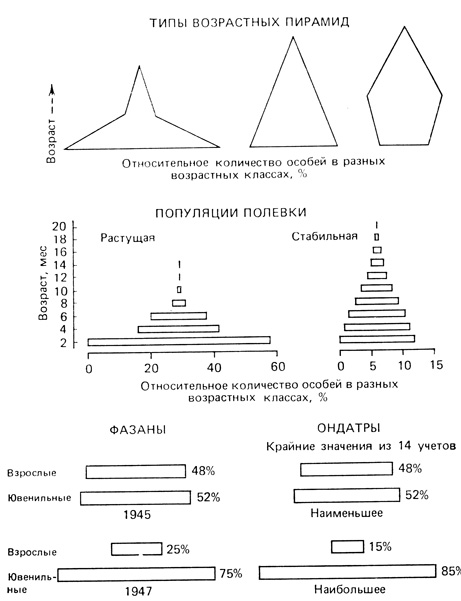
Рис. 6.5. Возрастные пирамиды. Вверху. Три типа возрастных пирамид, характеризующих популяции с высокой, умеренной и малой численностью (в %) молодых особей. В середине. Возрастные пирамиды лабораторных популяций полевки Microtus agrestis. Слева. Экспоненциальный рост популяции в нелимитирующей среде. Справа. Рождаемость и смертность равны. (Leslie, Ranson, 1940). Внизу. Крайние случаи соотношений численности ювенильных и взрослых особей у фазана в Северной Дакоте (Kimball, 1948) и ондатры на востоке США (Petrides, 1950)
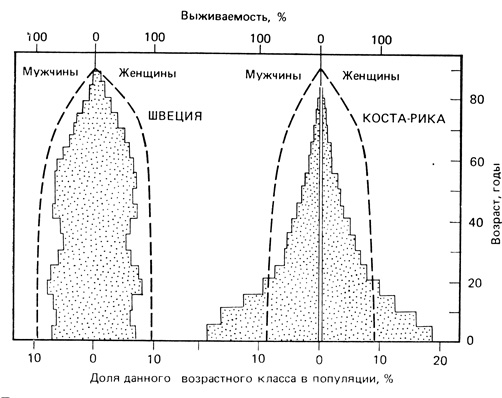
Рис. 6.6. Возрастная структура популяции человека, рассчитанная отдельно для мужчин и женщин в Швеции (1965 г.) и Коста-Рике (1963 г.). Население Швеции увеличивалось медленно, поэтому основание пирамиды узкое. Велик процент старших возрастных классов (40-60 лет). Расширение возрастного класса 15-23 года явилось следствием резкого увеличения рождаемости после второй мировой войны. Быстрый рост населения Коста-Рики сформировал пирамиду с широким основанием и высоким процентом населения моложе 25 лет. (Ricklefs, 1979, по данным Keyfitz, Fieger, 1968)
Упрощенно в популяции можно выделить три экологические возрастные группы: пререпродуктивную, репродуктивную и пострепродуктивную. Длительность этих возрастов по отношению к общей продолжительности жизни сильно варьирует у разных организмов. У современного человека три этих "возраста" приблизительно одинаковы, на каждый из них приходится около трети жизни. У первобытных людей пострепродуктивный период был намного короче. Для многих животных и растений характерен очень длительный пререпродуктивный период. У некоторых животных, в особенности у насекомых, пререпродуктивный период чрезвычайно длителен, репродуктивный период очень короткий, а пострепродуктивный период отсутствует совсем. Классические примеры - некоторые виды поденок (Ephemeridae) и 17-летняя цикада. У поденок личиночное развитие в воде занимает от года до нескольких лет, а во взрослом состоянии они живут всего несколько дней. У цикад необычайно продолжителен период развития (хотя и не обязательно 17 лет), но взрослые насекомые живут менее одного сезона.
У промысловых птиц и пушных зверей отношение сеголетков к старшим возрастным группам служит показателем тенденции изменения численности популяции. Это отношение определяют во время охотничьего сезона (осенью или зимой) по выборке из популяции, полученной охотниками или промысловиками. В общем высокая численность молоди по сравнению с численностью взрослых, как это видно из нижних схем на рис. 6.5, указывает на то, что сезон размножения сложился успешно; и если смертность молоди не будет чрезмерной, то можно ожидать увеличения популяции в следующем году. Наибольший процент молоди (85%) отмечался в популяции ондатры (рис. 6.5 внизу справа), которая несколько предшествующих лет подвергалась интенсивной промысловой нагрузке. Вызванное таким воздействием общее снижение численности популяции привело, по-видимому, к увеличению рождаемости у выживших особей, что указывает на гибкость, свойственную популяциям ондатры, - они быстро восстанавливают свою численность после периодов высокой смертности.
Явление, известное под названием "доминирование возрастного класса", часто наблюдается в популяциях рыб, для которых характерна высокая потенциальная рождаемость. Когда в результате чрезвычайно высокой выживаемости икры и личинок значительную часть популяции составляют особи определенного возрастного класса, в последующие несколько лет размножение подавляется. Классический пример этого - данные Порта по сельди в Северном море (рис. 6.7). Рыбы возрастного класса 1904 года доминируют в уловах с 1910 года (когда особи этой группы достигли 6-летнего возраста; к этому времени они уже достаточно крупны и эффективно вылавливаются промысловыми орудиями) до 1918 года (когда в 14-летнем возрасте они еще превосходили по численности рыб младших возрастных групп). Такая ситуация порождает иногда периодичность общих уловов, которые были велики в 1910 г. и снижались в последующие годы по мере снижения численности доминирующего возрастного класса до тех пор, пока его не заменили другие классы. Мы наблюдаем здесь нечто вроде компенсаторного механизма, когда высокая выживаемость с большой вероятностью сопровождается годами низкой выживаемости. Ихтиологи еще не пришли к единому мнению относительно того, какие внешние факторы обусловливают наблюдающийся время от времени необычный характер выживания.
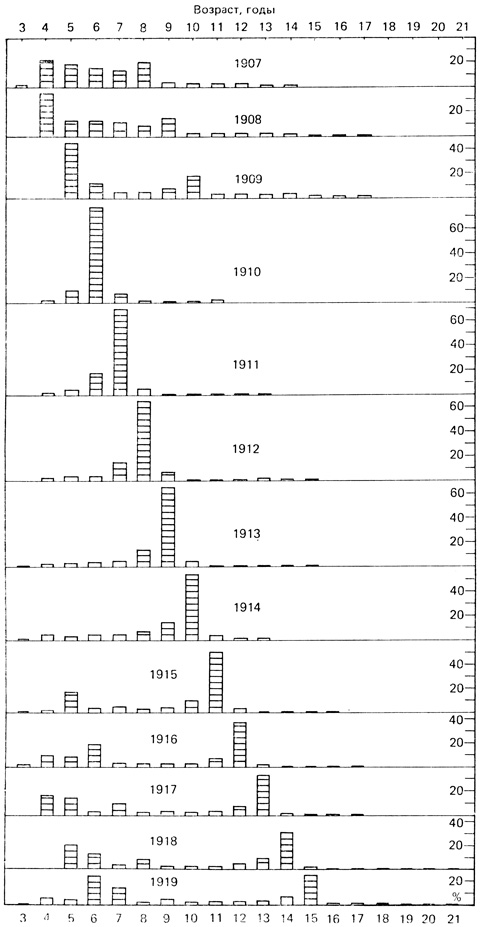
Рис. 6.7. Возрастное распределение сельди Северного моря в промысловых уловах за период с 1907 по 1919 г. иллюстрирует феномен доминирующего возрастного класса. Особи 1904 г. очень многочисленны и доминируют в популяции на протяжении многих лет. Рыбы моложе 5 лет не попадают в сети, поэтому класс 1904 г. не выявляется до 1909 г. Возраст рыб определяли по кольцам роста на чешуе, которые образуются ежегодно подобно годичным кольцам на деревьях. (По Hjort, 1926)
https://f-service.su услуга по замене фискального накопителя.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'