3. Речные экосистемы и поймы
Сколько рек на Земле и какова их общая протяженность? На этот вопрос ответить непросто. Ю. П. Беличенко и Л. Я. Полянинов (1976) утверждают, что общая протяженность рек только на территории СССР составляет 3 млн. км. Однако И. В. Попов (1977) в своей книге о речных руслах сообщает, что на территории СССР имеется около 3 млн. рек (!) общей протяженностью более 10 млн. км. По-видимому, И. В. Поповым учитывались даже весьма мелкие, постоянно действующие русловые водотоки длиной в несколько сот метров. Если исходить из данных И. В. Попова и учесть, что территория СССР занимает около 15% всей площади суши, то можно считать, что на Земле имеется около 20 млн. рек общей протяженностью 67 млн. км.
По данным П. Вейля (1977), относительная площадь бессточных районов, т. е. районов, не имеющих речного стока в Мировой океан, для суши в целом составляет около 33%, а для отдельных континентов она колеблется от 10% (в Северной Америке) до 6,4% (в Австралии). При этом лишь 800 тыс. км2 бессточных котловин лежит ниже уровня моря (Толстой, 1975).
Одним из крупнейших в мире бессточных районов, например, является Арало-Каспийский бассейн. Площадь Арало-Каспийской области замкнутого стока, включающей также бассейны озер Балхаш, Иссык-Куль и др., равна 5,45 млн. км2, что составляет 23,9% территории СССР (Энергетические ресурсы СССР. Гидроэнергетические ресурсы, 1967).
Территорию, подобную Арало-Каспийскому бассейну, уместно считать единой, хотя и открытой экосистемой. Эпитет "открытая" в данном случае совершенно не смягчает остроты постановки вопроса. В конце концов существуют ли вообще в природе абсолютно закрытые экосистемы, этакие "вещи в себе"? Любую из них, даже экспериментальную, при зрелом размышлении можно низвести в ранг открытой... Вопрос, следовательно, состоит лишь в том, какие же преимущества дает, а точнее, может дать подход, при котором река и ее водосборный бассейн рассматривались бы как элементы единой и целостной экосистемы?
Собственно, балансовые гидрологические расчеты основываются всегда именно на таком подходе. Экологи же привыкли в данном случае оперировать чрезвычайно мелкими категориями: экосистемы устьевого участка реки, лотические сообщества (сообщества перекатов и плёсов), экосистемы ручья, экосистемы низкой поймы, экосистемы свежих аллювиальных наносов и т. п. Ю. Одум (1975) справедливо подчеркивает, что, "учитывая интересы человека, минимальной единицей экосистемы следует считать не один водоем, а всю площадь водосбора... Причины загрязнения вод и способы борьбы с ним не удается обнаружить, если смотреть лишь на воду; наши водные ресурсы страдают из-за плохого хозяйствования на всей площади водосбора, который и должен рассматриваться в качестве хозяйственной единицы" (с. 26). Специалисты из Северо-Кавказского научного центра высшей школы на своей модели экосистемы Азовского моря убедительно продемонстрировали жизнеспособность и актуальность высказанных идей. По-видимому, в условиях нашей огромной страны имеет смысл постепенно приводить границы административных районов и территориальных хозяйственных единиц в соответствие с границами водосборных бассейнов. Целесообразность подобных реформ была достаточно обоснована еще А. И. Воейковым и В. В. Докучаевым.
Каждый бассейн, каждый водосбор уникален, однако именно поэтому и надо попытаться прежде всего расположить все их многообразие хотя бы в самую примитивную схему. Принципы построения классификации могут быть самые разнообразные, но нам следует предусмотреть возможность экологической оценки каждой из выделенных категорий водосборов, соблюдая при этом определенную преемственность между отдельными классификационными единицами.
Прежде всего мы должны выделить в самостоятельный тип водосборы, не имеющие речного стока в Мировой океан и не имеющие также внутреннего открытого водоприемника. С точки зрения не только экологии такая система является наиболее замкнутой: она ближе всего стоит к типу идеальных и гипотетических закрытых экосистем. В пределах этого типа можно выделить по крайней мере два подтипа: системы, не имеющие русловой гидросети, и системы, имеющие русловую, хотя бы и периодически действующую гидросеть. Рассмотрим, например, водосбор пустыни Рамлат-эс-Сабъатайн в Йеменской Арабской Республике, относящийся именно к первому типу и ко второму подтипу водосборов. Нет сомнения, что вся эта система имела некогда прямой сток в Аденский залив Аравийского моря, а еще раньше и сама в своей пониженной части была заливом этого моря. В данном случае мы имеем дело с реликтовой, деградировавшей и чрезвычайно древней гидросетью.
Далее мы можем выделить в самостоятельный тип водосборы, также не имеющие речного стока в Мировой океан, но имеющие открытый сток во внутриконтинентальные водоемы. В особые подтипы при этом можно выделить водосборы, имеющие сток в соленые озера и в пресные. Надо полагать, что в большинстве случаев водосборы с соленым "резервуаром" более древние, реликтовые. Таков, например, бассейн соленого оз. Ван в Турции. У нас в стране много систем подобного подтипа (Волго-Каспийская система, системы Аральского моря и оз. Балхаш), но все они чрезвычайно изменены человеческой деятельностью.
Водосборы, имеющие прямой речной (русловой) сток в Мировой океан, вполне естественно подразделить на три подтипа: бассейны, имеющие сток непосредственно в океан (в море); водосборы, имеющие сток в эстуарий, т. е. в губу, в залив Мирового океана; наконец, самые распространенные на земном шаре экосистемы, имеющие речной сток в речную гидросеть более высокого порядка.
Водосборы выделенных типов существенно различаются между собой по географическому распространению, по происхождению, по ритмике стока и поведению вещества (косного и биогенного). Нет сомнения, например, что, соорудив водохранилище в бассейне последнего типа, мы переводим часть его водосбора, лежащую выше водохранилища, в режим предыдущего типа. Возникает справедливый вопрос: в чем состоят принципиальные экологические различия между водосборами рассматриваемых типов независимо от их известной географической изменчивости? Можно конкретизировать вопрос, например, таким образом: существуют ли (известны ли) сугубо биологические различия между бассейнами рек Терека и Кубани исключительно из-за того, что Терек впадает во внутриконтинентальный водоем, а Кубань, хотя и опосредованно, имеет связь с Мировым океаном? Уровень Каспия, как известно, лежит на 28 м ниже уровня Черного моря.
Вполне очевидно, что первым каналом связи "снизу - вверх" для водосбора является базис эрозии. Повышение-понижение его через русловые процессы в гидросети несомненно отражается на состоянии всего водосбора, и в первую очередь на сукцессионных превращениях биоценозов, тяготеющих к водным коммуникациям. Биологические эффекты подобного рода могут быть чрезвычайно тонкими и различаться лишь при ювелирном геоботаническом исследовании. Здесь напрашивается определенная аналогия с проблемами, возникающими, например, при оценке биологических эффектов электромагнитных полей ЛЭП. И в том и в другом случае особое значение приобретают индикационные аспекты геоботаники и биогеоценологии: подбор тест-объектов; выбор признаков биосистем, чувствительных к данному фактору; совершенствование методов сбора и обработки материала массовых экологических наблюдений.
Вторым каналом связи между водоприемником и водосбором являются гидробионты. Их поведение с этой точки зрения, за исключением миграционных передвижений рыб, изучено еще недостаточно. Надо полагать, что значительная роль в перемещении низших гидробионтов (бактерий, водорослей, грибов, беспозвоночных) от устья реки по направлению к верховьям принадлежит наземным животным и особенно птицам. Со времен Дарвина и до наших дней биологи в своих теоретических представлениях о биологических коммуникациях громадное значение придают нередко лишь одной стороне дела, а именно фактору изоляции. В гидробиологии, однако, в силу чрезвычайной динамичности субстрата основополагающим во многих ситуациях может быть как раз фактор взаимодействия между, казалось бы, заведомо разобщенным в пространственно-временном отношении биологическим материалом. Рассмотрим более подробно одно из удивительнейших явлений отмеченного порядка.
Колебания базисов эрозии и их асинхронность наряду с локальными геоморфологическими процессами в смежных речных бассейнах приводят к речным перехватам и переливаниям рек (Болиг, 1956). При этом отдельные участки водосбора одной реки вместе с их биоценозами становятся принадлежностью другой реки, иногда весьма отличающейся по своим географическим и биологическим особенностям. По сообщению Е. В. Ястребова (1961), например, подобные явления, можно сказать, в классическом виде широко распространены на Северном Урале, во взаимоотношениях между бассейнами рек Вишеры и Лозьвы. Притоки Лозьвы с их более низкими долинами и базисами эрозии растут в длину в западном направлении, постепенно перехватывая сток меридиональных отрезков рек и ручьев западного макросклона Уральского хребта. При этом генеральный водораздел Европа - Азия постепенно смещается к западу, отдельные участки территории вместе с соответствующими биоценозами отходят к бассейну Оби. На рис. 2 показана принципиальная схема этого процесса, однако в ней не учтено, что в связи с изменением водоносности на рассматриваемых участках рек должны существенно измениться и русловые процессы. Разумеется, при специальном подходе можно найти немало способов абсолютной датировки и определения скоростей прохождения отдельных фаз этих процессов. С биологической точки зрения наибольший интерес в данном случае представляет то обстоятельство, что перехват части водосборов западного макросклона Урала реками восточного склона сопровождался своего рода "дрейфом" элементов европейского флористического и фаунистического комплексов на восток. Нечто подобное, только в более широких масштабах, может происходить и при перебросках стока рек.
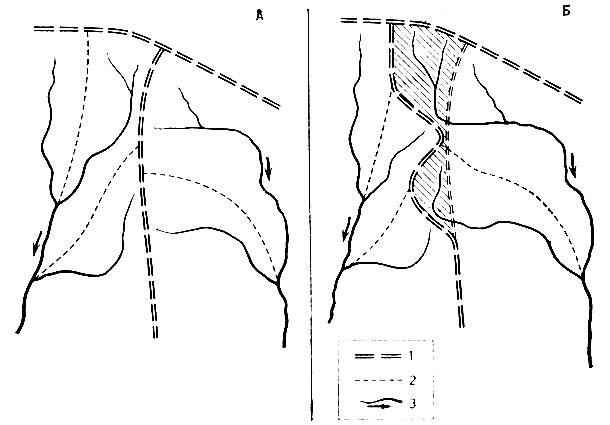
Рис. 2. Принцип перехвата стока одной реки другой: 1 - границы генеральных водосборов; 2 - границы второстепенных водосборов; 3 - реки
При прочих равных условиях большое экологическое значение имеет относительное распределение территории водосборного бассейна по категориям сточных, транзитных и аккумулятивных ландшафтов, которое можно назвать "архитектоникой" бассейна. Это положение иллюстрируется в самом общем виде, на широком географическом фоне. Действительно, если принять площадь аккумулятивных ландшафтов в бассейне и вообще на какой-либо территории за константу, а сумму площадей сточных и транзитных ландшафтов - за "единицу" (1,0), то, исходя из известных зональных показателей среднегодового объема осадков и испаряемости, можно рассчитать объемы стока с водосборов в магистральные водотоки при различных соотношениях сточных и транзитных площадей. В табл. 2 приведены результаты такого расчета для нескольких ландшафтно-географических зон. Расчет сделан для четырех вариантов соотношения площадей сточных и транзитных ландшафтов в бассейне (0,8:0,2; 0,6:0,4; 0,6:0,4; 0,2:0,8).
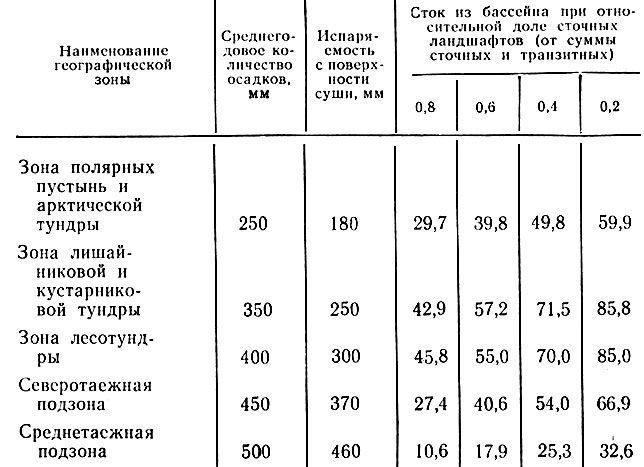
Таблица 2. Расчет среднегодового объема стока, поступающего в основную гидросеть с водосбора, в зависимости от архитектоники бассейна (соотношения сточных и транзитных площадей), мм/год (Плотников, 1984)
Как видно, влияние этого фактора весьма существенно и отличается определенными зональными особенностями. Этот абстрактный расчет представляет отнюдь не схоластический интерес. В связи с водохозяйственным строительством (плотин, каналов) и прочими видами промышленной деятельности, особенно на равнинных территориях, мы невольно переводим некоторые площади из категории сточных в категорию транзитных и наоборот, автоматически нарушая тем самым исторически сложившийся водный баланс отдельных бассейнов. Рассуждая аналогичным образом конкретно о короткой истории экосистемы бассейна р. Хадытаяха (Южный Ямал), было показано (Плотников, 1984), что доля транзитных ландшафтов в нем неуклонно увеличивалась за счет денудационного разрушения поверхности водоразделов, а значит, и приток воды с водосбора в долину реки при прочих равных условиях увеличивался (и увеличивается). Иными словами, было доказано, что формирование долинного биогеоценологического комплекса данной экосистемы протекало на фоне растущей обводненности, если даже атмосферное увлажнение и не увеличивалось. Экологическое значение подобных обстоятельств трудно переоценить.
Заманчивую перспективу для высокой формализации гидрографических и геоморфологических, а впоследствии и экологических представлений дает наблюдение И. Н. Степанова (1982): вода, стекающая с гор, способствует выработке на прилегающей равнине рельефа, антисимметричного рельефу горной системы. И в данном случае весьма наглядным образом подтверждается гениальная догадка В. И. Вернадского об универсальном методологическом значении явлений симметрии-дисимметрии в неживой и живой природе.
Аллювиальные наносы рек, как новейшие в геологическом отношении естественные субстраты, привлекают особое внимание экологов и геоботаников, исследующих смены растительного покрова. Замечательной особенностью аллювиальных отложений рек является их разновозрастность в пределах отдельных структурных элементов поймы (аллювиальных сегментов, островов), что определяет одновременное присутствие на них растительных сообществ, находящихся на разных стадиях индивидуального развития (сукцессии). Это обстоятельство позволяет из доступной наблюдению статики процесса аллювиальной сукцессии вывести его динамику, на что указывали в свое время К. Н. Игошина (1927) и Б. Н. Городков (1946). Интересный в этой связи анализ идей зарубежного поймоведения проделан Б. М. Маркиным и Н. Г. Ишбулатовой (1967).
Наиболее полные схемы процесса смен растительности на речных наносах даны Б. Н. Городковым (1946) для севера лесной зоны Западной Сибири и В. Шелфордом (1954) для нижнего течения Миссисипи (Shelfod, 1954). Оригинальные схемы или фрагменты схем составлены для аллювиев Камы и Чусовой (Игошина, 1927), долины Печоры (Самбук, 1930), верхней Оби (Бокк, 1966), Бурей (Нечаев, 1967), верховьев Вишеры (Плотников, 1968, 1970), некоторых рек Аляски (Drury, 1956; Bliss, Cantlon, 1957). Обобщенная схема аллювиальной сукцессии представляется в следующем виде:
1. Стадия пионерных растительных группировок на свежих аллювиальных наносах, отмеченная преобладанием ползучекорневищных трав. Как показал А. П. Нечаев (1967), семена ивовых имеют специальные приспособления для заселения субстрата уже на этой стадии.
2. Стадия заливного луга; на горных отрезках рек обычно весьма быстротечная, во многих случаях длительно поддерживается хозяйственной деятельностью человека.
3. Стадия заливных кустарников, преимущественно ивовых.
4. Стадия первичного лиственного леса.
5. Стадия первичного хвойного леса.
6. Стадия субклимакса, т. е. биоценоза, близкого по своей структурно-функциональной организации к зональному типу ценозов, но не сменяемого последним в силу ощутимого влияния гидрологического и микроклиматического факторов речной долины.
Длительность отдельных стадий сукцессии и общая продолжительность полной серии смен растительности на речном аллювии зависят от многих обстоятельств и могут быть вычислены несколькими способами. Наиболее популярные способы основаны на детальной геоботанической съемке какого-либо конкретного острова или аллювиального сегмента и последующем ретроспективном хронологическом анализе (абсолютной датировке) отдельных разновозрастных элементов речных отложений и растительного покрова на них. Особую ценность при этом представляют исследования годичных приростов деревьев и кустарников. Датировки осуществляются, как правило, с весьма высокой точностью, а затем распространяются на определенные отрезки речной долины. Разумеется, чрезвычайно ценные результаты можно получить при анализе серии разновременных лоцманских карт одной и той же реки.
Нами разработан своего рода экспресс-метод определения скорости аллювиальных сукцессий, основанный на использовании материалов разовой аэрофотосъемки местности (Плотников, 1968, 1970). Для этого на определенном, генетически однородном участке реки (следует особо выделять по крайней мере верховья, среднее течение и низовья реки) путем дешифрирования аэрофотоснимков собираются характеристики как можно большего количества островов или сегментов. На каждом из островов выделяются зоны, соответствующие отдельным стадиям аллювиальной сукцессии (песчано-галечные косы с пионерными группировками растений, кустарниковые заросли, первичный лиственный лес, темнохвойный лес). Выбираются признаки, надежно дешифрируемые на снимках даже весьма низкого качества. На цветных и спектрозональных крупномасштабных снимках можно отдешифрировать множество промежуточных элементов сукцессии. По наличию или отсутствию отдельных зон, соотношению их площадей, по размерам и конфигурации островов их можно разбить на несколько групп, соответствующих гипотетическому генетическому ряду. Далее составляется и решается система уравнений, в которых пространственные параметры островов и количественные соотношения аллювиальных зон на них ставятся в пропорциональную зависимость от времени их существования. При некоторых специфических в каждом конкретном случае допущениях таким путем удается установить общую длительность сукцессии и ее отдельных этапов или стадий.
Наблюдения, выполненные самыми различными способами, показывают, что скорость сукцессии уменьшается, а ее продолжительность соответственно увеличивается по направлению от истоков реки к ее низовьям. Для крупнейших рек умеренного пояса северного полушария, таких, как Обь и ее притоки, Кама, Печора, длительность полного цикла аллювиальной сукцессии приближенно можно считать равной 200 - 250 годам в верхнем, 400 - 500 годам в среднем и 600 - 1000 годам в нижнем течении. Разумеется, самостоятельное значение в этом случае имеют и зональные климатические особенности, но выяснить его можно лишь на достаточно широком географическом фоне.
"Наши реки имеют некоторые особенности сравнительно с реками Западной Европы, всего лучше изученными до сих пор. Две главные из этих особенностей: малое падение и правильность половодья весной", - писал А. И. Воейков еще в 1888 г. (Воейков, 1963, с. 193). Как было показано выше, скорость аллювиальных сукцессий практически прямо пропорциональна величине падения реки. Из-за этого мы располагаем весьма внушительными ресурсами речных пойм. По данным П. А. Худякова (1976), площадь пойм составляет 2,5% от всей площади СССР. По его же расчетам, поймы рек производят не менее 30% всей биопродукции континентальной части земного шара. В масштабах всей планеты, по сообщению А. В. Лапо (1979), поймы занимают меньше 1% поверхности, а производят около 10% живого вещества. Таким образом, биологическая продуктивность пойменных экосистем на порядок величин превосходит продуктивность всех других континентальных ландшафтов. Специфические особенности пойм (по П. А. Худякову) следующие:
1. Пойма, как аккумулятор питательных веществ, сама и гарантирует, и регулирует свое плодородие.
2. В паводково-пойменном экологическом цикле складывается исключительно благоприятный газово-термический режим.
3. В нем смягчена суровость морозного периода.
4. Пойма представляет собой арену регулярных миграций животных.
5. Пойма - место переживания зимы для многих видов животных.
6. Сложная архитектоника пойм является причиной исключительного разнообразия их растительного и животного населения.
7. Пойма хорошо и регулярно промывается от всевозможных "шлаков".
8. Поймы до сих пор сохраняются в относительно не нарушенном состоянии (если не учитывать грандиозные затопления, спрямления русел рек и т. д.).
К этому перечню следует добавить, что пойма - это убежище для растений и животных от многих неблагоприятных, так называемых исторических факторов (урбанизаций, войн и пр.). Особую ценность представляют пойменные луга. Характеризуя их, А. Н. Тюрюканов, И. Т. Кузьменко и М. Т. Павлова (1971) утверждают, в частности, что "качеству биопродукции пойменных лугов следует отдать предпочтение перед ее количеством" (с. 50). Тенденция превращать пойменные луга в пашню чрезвычайно неблагоприятна еще и потому, что "для смыва почвы толщиной 15 см на пашне требуется всего 15 лет, а на лугу - 3500 лет" (Попов, 1977, с. 83). Даже в Субарктике отдельные типы пойменных лугов могут производить в год до 150 ц/га надземной биомассы в воздушно-сухом весе (Пешкова, 1977). По-видимому, проблема рационального использования пойменных земель будет решена тогда, когда агрономы, животноводы и экономисты будут давать товарную оценку каждому квадратному метру поймы, изымаемому для каких-либо нужд из естественного биологического оборота.
Специфическим элементом пойм равнинных сибирских рек являются так называемые "сора" или "туманы" - мелководные, то промерзающие, то обсыхающие озеровидные разливы рек преимущественно в их устьевых участках (Вагильский Туман на р. Вагиль, Пелымский Туман на р. Пелым; Турсунский, Шаимский Туманы и Кондинский Сор на Конде, Харбейский, Веленгамский и Воронковский Сора на Оби и многие другие). Известно, что это нерестилище и нагульные пастбища ценнейших пород рыб, а также великолепные охотничьи угодья. Они же являются и регуляторами ледовой обстановки на реках... Но каков генезис этих водоемов, какова их связь с новейшей тектоникой Западной Сибири, наконец, каково их будущее в связи, например, с эксплуатацией знаменитых нефтяных и газовых месторождений? Эти и десятки других, более специальных вопросов ждут своего незамедлительного разрешения.
Анализируя современные представления о влиянии лесохозяйственных и лесомелиоративных мероприятий на речной сток, иногда высказываются лишь некоторые общие, "гарантийные" соображения: "...при сокращении лесопокрытой площади всей лесной зоны примерно на 15% водный режим рек заметно ухудшится, что выразится прежде всего в ослаблении подземного питания из-за заиления источников в результате возросшей эрозии и из-за нарушения водного баланса" (Идзон, 1974, с. 72). Но что такое лесная зона? На территории нашей страны она слишком обширна и неравномерно освоена, чтобы рассматриваться в качестве единой хозяйственной единицы. Лесная зона - это прежде всего своего рода конгломерат водосборных бассейнов различной облесенности. Концепция водосбора как элементарной водохозяйственной и естественноисторической категории представляется в данном случае весьма конструктивной. Действительно, расположив, например, все водосборы лесной зоны в порядке уменьшения их лесистости, мы одновременно получим ряд рек, в котором наверняка возрастает амплитуда внутригодовых колебаний стока. Далее следует установить более тонкие детали, а именно как зависят относительные параметры стока от видового состава и возрастной структуры лесов водосбора. Наконец, особый интерес представляет выяснение зависимости параметров стока от соотношения облесенности истоков реки, облесенности транзитных отрезков реки и ее устьевой части. Любопытные результаты по исследованию влияния залесенности, заболоченности и озерности территории водосбора на некоторые характеристики стока для отдельных районов Западной Сибири приводятся в книге "Закономерности и прогнозирование природных явлений" (1980). Данные о зависимости параметров речного стока от лесистости водосборных бассейнов приведены И. С. Сафаровым (1982) для горных экосистем Азербайджанской ССР.
Как видим, требования экологического подхода отнюдь не сложны, и решение этих вопросов даст нам в руки надежную научную основу для планирования водохозяйственных мероприятий. Именно пересмотр официальных форм учета лесного фонда нашей страны, основанных на административном и ведомственном соподчинении территории, и приближение содержания и структуры учета лесного фонда к рассмотрению водосбора как элементарной статистической единицы гарантирует понимание функциональной природы лесов страны по отношению к ее водному хозяйству.
С экологической точки зрения очень важен учет объемов и "поведения" теплового стока рек, особенно в северных районах страны. Тепловым стоком называют количество тепла, переносимое рекой через какое-либо ее сечение за единицу времени (Одрова, 1980). Тепловой сток 0 за время Т вычисляется по формуле
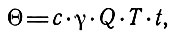
где t - средняя температура воды, Q - расход воды (объем воды, протекающий за секунду через данное сечение реки), c - теплоемкость воды и γ - ее удельный вес.
Из формулы следует, что величина теплового стока зависит главным образом от температуры и расхода воды, которые подвержены сезонным и многолетним колебаниям и доступны регулированию. Максимум теплового стока рек приходится на летние месяцы. Это и определяет его огромное экологическое значение.
В мировой практике сложилось и наполняется все новым содержанием понятие "международная река" (Чичварин, 1970). Вполне очевидно, что концепция водосборного бассейна как единой и целостной экосистемы приобретает в данном случае международно-правовое и экономическое значение.
В последние годы общественность начала проявлять интерес к проблемам малых рек. Известный знаток речных систем С. Л. Вендров определяет их следующим образом: "К малым обычно относят реки, у которых площадь водосбора не более 2 тыс. кв. км или же у которых длина не более 100 км независимо от площади водосбора. Критерии эти, конечно, весьма условны" (Наука и жизнь, 1983, № 9, с. 2). Малые реки у нас в стране составляют около 99% от их общего числа, а их суммарная длина достигает 92% от общей протяженности рек Советского Союза. Комментарии к этим данным поистине излишни.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'