Объяснения и примеры
Описательные исследования сукцессий на песчаных дюнах, в степях, лесах, на морских берегах или в других местах наряду с более новыми функциональными исследованиями позволили построить ряд теорий, частично объясняющих процесс развития и его причины. Г. Т. Одум и Пинкертон (Н. Т. Odum, Pinkerton, 1955), опираясь на сформулированный Лоткой (Lotka, 1925) "закон максимума энергии в биологических системах", впервые указали на то, что сукцессия связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленной на поддержание (дыхание) системы, по мере того, как накапливаются биомасса и органическое вещество. Позднее Маргалеф (Margalef, 1963, 1968) подкрепил эту концепцию о биоэнергетических основах сукцессии фактическими данными и расширил ее. В последние 15 лет интенсивно обсуждается роль, которую играют межпопуляционные взаимодействия в формировании последовательности смены видов, характерном признаке экологической сукцессии. (См. соответствующие обзоры Connell, Slayter, 1977; McIntosh, 1980.)
Изменения основных структурных и функциональных характеристик аутогенной сукцессии перечислены в табл. 8.1. Для удобства обсуждения двадцать признаков экологических систем разбиты на пять групп. Начальные и конечные стадии развития характеризуются различными тенденциями. Абсолютная величина изменений, скорость изменений и время, необходимое для достижения стационарного состояния, могут варьировать не только различных климатических и физико-географических ситуациях, но и для разных признаков экосистемы в одной и той же физической среде. При наличии хороших исходных данных кривые скорости изменений имеют обычно выпуклую форму (причем в начале изменения происходят быстрее). Известны также двувершинные или периодические кривые.
Таблица 8.1. Тенденции изменения основных характеристик экосистемы, которых следует ожидать в ходе аутогенной сукцессииЭнергетика экосистемы 1. Возрастают биомасса (В) и количество органического детрита 2. Возрастает валовая продукция (Р) за счет первичной; вторичная продукция изменяется мало 3. Уменьшается чистая продукция 4. Увеличивается дыхание (R) 5. Соотношение P/R приближается к единице (равновесию) 6. Соотношение В/P возрастает (P/В соответственно уменьшается) Круговороты биогенных элементов 7. Круговороты элементов становятся все более замкнутыми 8. Увеличиваются время оборота и запас важных элементов 9. Возрастает коэффициент цикличности (возобновление/вход) 10. Удерживается и сохраняется больше биогенных элементов* Виды и структура сообщества 11. Меняется видовой состав сообщества (флористические и фаунистические эстафеты) 12. Возрастает богатство как компонент разнообразия 13. Возрастает выровненность как компонент разнообразия 14. r-Стратеги в широких масштабах замещаются K-стратегами 15. Усложняются и удлиняются жизненные циклы 16. Увеличивается величина организмов и (или) их стадий расселения (семян, молоди и т. д.) 17. В значительной степени развивается взаимовыгодный симбиоз* Стабильность 18. Возрастает резистентная устойчивость* 19. Снижается упругая устойчивость* Общая стратегия 20. Возрастает эффективность использования энергии и биогенных элементов*
* (Тенденции обоснованы теоретически и еще не подтверждены в полевых условиях.)
Перечисленные в табл. 8.1 тенденции наблюдаются в том случае, если в сообществе преобладают внутренние, аутогенные, процессы. Влияние внешних, аллогенных, возмущений может повернуть вспять или иным образом изменить эти тенденции развития. Эти вопросы мы обсудим позднее.
Биоэнергетика развития экосистемы. Биоэнергетику экосистемы отражают первые шесть пунктов из табл. 8.1. На ранних стадиях автотрофной сукцессии в среде, лишенной органического вещества, скорость образования первичной продукции, или общий (валовой) фотосинтез (P), превышает скорость дыхания сообщества (R), так что соотношение P/R, как правило, больше 1. В особых условиях, когда сукцессия протекает в богатой органикой среде (например, в очистном отстойнике), соотношение P/R бывает меньше 1. Такую среду первыми заселяют бактерии и другие гетеротрофные организмы, поэтому сукцессия называется гетеротрофной. Однако в обоих случаях, согласно теории, величина P/R приближается к единице по мере развития сукцессии. Иными словами, в зрелой, или климаксной, экосистеме наблюдается тенденция к равновесию между связанной энергией и энергией, затрачиваемой на поддержание биомассы (т. е. суммарным дыханием сообщества). Таким образом, отношение P/R является функциональным показателем относительной зрелости экосистемы. Отношения P/R для ряда хорошо изученных экосистем приведены на рис. 8.1, А, где указано также направление автотрофной и гетеротрофной сукцессий.
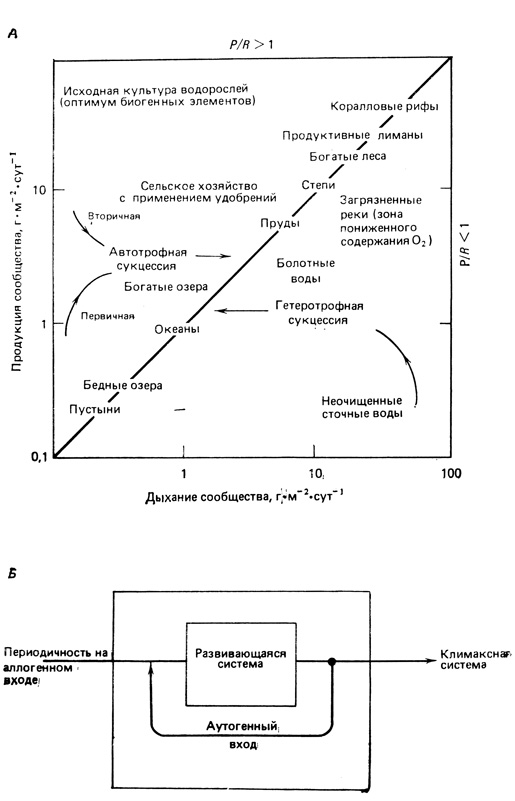
Рис. 8.1. А. Положение разных сообществ в системе классификации, основанной на метаболизме сообщества. Слева от диагональной линии валовая продукция (Р) превышает дыхание (R); P/R больше единицы = автотрофность. Справа от диагонали - обратная ситуация: P/R меньше единицы = гетеротрофность. В последнем случае сообщества живут за счет органического вещества, получаемого извне, или за счет его предварительного запасания или накопления. Стрелками показано направление автотрофной и гетеротрофной сукцессий. Сообщества, расположенные вдоль диагонали, в среднем за год потребляют примерно столько, сколько создают, и их можно считать метаболическими климаксами. (Из Н. Т. Odum, 1956.) Б. Общесистемная модель развития экосистемы
Пока P больше R, в системе будут накапливаться органическое вещество и биомасса (В), в результате чего отношения В/P, В/R или В/E (где E = P + R) будут увеличиваться или соответственно отношение P/В будет уменьшаться. Напомним, что эти отношения уже обсуждались в связи с концепцией термодинамической упорядоченности (т. 1, с. 109). Теоретически, следовательно, урожай на корню, поддерживаемый доступным потоком энергии (E), достигает максимума на зрелых, или климаксных, стадиях. Как следствие этого, чистая продукция сообщества (или урожай за годовой цикл) велика на ранних стадиях и мала или равна нулю в зрелой системе.
Сравнение сукцессии в лабораторном микрокосме и в лесу. Биоэнергетические изменения легко наблюдать, вызвав сукцессию в экспериментальных лабораторных микроэкосистемах, происходящих от природных систем; такие микроэкосистемы были описаны в гл. 2 (см. рис. 2.17, В). На рис. 8.2 общее течение 100-дневной автотрофной сукцессии в микрокосме (данные Cooke, 1967) сравнивается с гипотетической моделью 100-летней сукцессии леса (данные Kira, Shidei, 1967). В течение первых 40-60 дней типичного эксперимента с микрокосмом дневная чистая продукция (Р) превосходит дыхание ночью (R), поэтому в системе накапливается биомасса (В). После начального периода "цветения" примерно к 30-му дню скорости обоих процессов начинают снижаться и на 60-80-й день становятся приблизительно равными. По мере приближения к стационарному состоянию отношение В/P (в граммах суточной продукции углерода, поддерживающих 1 г углерода биомассы) возрастает от менее 20 до более 100. В климаксном сообществе не только сбалансирован автотрофный и гетеротрофный метаболизм, но крупные органические структуры поддерживаются невысокой суточной продукцией и дыханием. Меняется и относительное обилие видов, так что в конце 100-дневной сукцессии доминирующими становятся другие бактерии, водоросли, простейшие и мелкие ракообразные, а не те, которые доминировали вначале (Gorden et al., 1969).
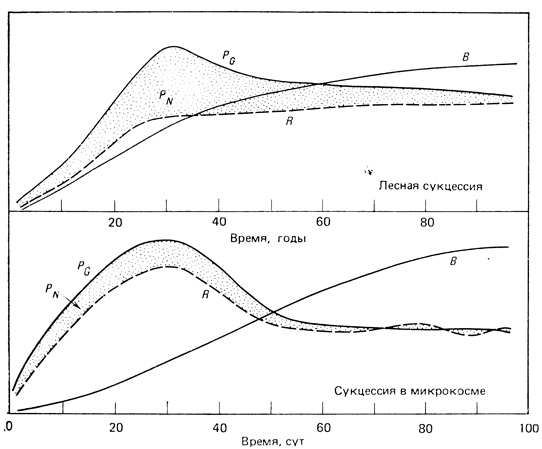
Рис. 8.2. Сравнение энергетики развития экосистем леса (из Kira, Shidei, 1967) и лабораторного микрокосма (из Cooke, 1967). PG - валовая продукция; PN - чистая продукция; R - суммарное дыхание сообщества; В - суммарная биомасса
В проточном лабораторном микрокосме, где культуральная среда медленно протекает через контейнер с организмами, имитируя вход и выход среды для воды и питательных веществ, сукцессия замедленна; Для накопления биомассы и приближения P/R к 1 требуется в 2-3 с лишним раза больше времени, чем в закрытом микрокосме (Hendrix et al., 1981).
Непосредственная экстраполяция данных, полученных для лабораторного микрокосма, на природные системы может быть и не вполне правомерна, поскольку в первом случае в системе существуют только мелкие организмы с простыми жизненными циклами и неизбежно снижено видовое и химическое разнообразие. Тем не менее в микрокосме можно наблюдать такие же основные тенденции, которые характерны для сукцессии на суше (рис. 8.2, верхний график) и в крупных водоемах. Сезонные сукцессии часто происходят по той же самой схеме: вслед за "цветением" в начале сезона, которое характеризуется быстрым ростом нескольких доминирующих видов, к концу сезона устанавливается высокое отношение B/Р, увеличиваются разнообразие и относительная, хотя и временная, устойчивость, как это установлено в терминах Р и R (Margalef, 1963). В открытых системах на зрелых стадиях понижения общей, или валовой, продукции, наблюдаемой в пространственно ограниченном микрокосме, может и не происходить, но общая схема биоэнергетических изменений в последнем, судя по всему, достаточно хорошо имитирует природу.
Интересно также отметить, что пик чистой первичной продукции (PN) соответствующий максимально возможному урожаю, приходится на 30-й день в микрокосме и 30-й год в лесу. Короткоцикличное лесное хозяйство ориентировано на пик PN, который во многих случаях повторяется через 20-40 лет.
Аллогенные и аутогенные воздействия. До сих пор мы рассматривали изменения, происходящие в результате процессов, протекающих в самой экосистеме. Привнесенные вещества или энергия, геологические силы, шторма, антропогенные нарушения и т. п. могут изменить, остановить или повернуть вспять тенденции, указанные в табл. 8.1 (так обычно и бывает на самом деле). Эвтрофикация озера, например (неважно - естественная или антропогенная), происходит в результате поступления в озеро биогенных элементов и почвы извне, т. е. с водосбора. Это равнозначно добавлению биогенов в лабораторную микроэкосистему или удобрению поля; в сукцессионном отношении такая система отбрасывается назад, к более молодому состоянию. Аллогенная сукцессия такого типа во многих аспектах обратна аутогенной сукцессии. Если влияние аллогенных процессов намного сильнее влияния аутогенных процессов, как во многих прудах и малых озерах, то экосистема не только не в состоянии стабилизироваться, но и "угасает", т. е. заполняется органическим веществом и осадками, превращаясь в болото или наземное сообщество. Такова неизбежная судьба искусственных озер, берега которых подвержены ускоряющейся искусственной эрозии.
Исследования озерных осадков (Mackereth, 1965; Cowgill, Hutchinson, 1964; Harrison, 1962) наряду с теоретическими соображениями указывают, что озера могут развиваться и действительно развиваются в сторону более олиготрофного (менее обогащенного) состояния, если поток биогенных элементов с водосбора замедляется или прекращается. Таким образом, есть надежда, что вызывающую беспокойство антропогенную эвтрофикацию, которая ухудшает качество воды и укорачивает жизнь водоема, можно повернуть вспять, если удастся сильно уменьшить приток биогенов с водосбора. Примером служит описанное Эдмондсоном (Edmondson, 1968, 1979) "выздоровление" озера Вашингтон в Сиэтле. В течение 20 лет очищенные, богатые биогенами сточные воды сливались в озеро, которое становилось все более мутным, и в нем все чаще наблюдались обильные "цветения" водорослей. В результате призывов общественности сточные воды отвели от озера, и оно быстро вернулось к более олиготрофному состоянию (вода стала чище, и прекратились "цветения" водорослей).
Взаимодействие внешних и внутренних сил можно выразить ври помощи обобщенной системной модели (рис. 8.1, Б). Впервые модель такого типа была приведена на рис. 1.4. Аутогенные силы изображены как внутренний вход или обратная связь, которая теоретически способствует сдвигу системы к своего рода равновесному состоянию. Аллогенные силы изображены как периодическое внешнее возмущение на входе, которое возвращает назад или иным способом меняет траекторию развития системы.
В тех системах, которые развиваются длительное время, как это бывает, например, при зарастании лесом голой почвы, сукцессионный процесс подвержен влиянию периодических возмущений, особенно в изменчивой среде умеренной зоны. Оливер и Стефенс (Oliver, Stefens, 1977) описали изменения растительного покрова на небольшой площади Гарвардского леса (Массачусетс). Растительность этой территории 14 раз подвергалась воздействиям природных и антропогенных стрессов разной силы с нерегулярными промежутками за период с 1803 по 1952 г. Имеются также данные о том, что до 1803 г. здесь пронеслось два урагана и возник один пожар. Слабые воздействия не вызывают появления новых видов деревьев, но часто позволяют видам, уже представленным в нижнем ярусе (черная береза, красный клен, тсуга), достигать полога. Крупномасштабные возмущения, такие, как ураган или сильный пожар, создают открытые пространства, прогалы, в которые вторгаются виды, свойственные ранним стадиям сукцессии (например, белая береза или пенсильванская черемуха). В таких прогалах развивается также молодая поросль из находившихся в почве семян и проростков. Северный дуб часто заполняет таким образом прогалы и через несколько десятилетий начинает доминировать в пологе. Смена видов и сукцессия в лесных прогалах была названа Брэем (Bray, 1956) "сукцессией фазы заполнения пробела". Оливер и Стефенс на основе полученных ими данных пришли к выводу, что существующий видовой состав леса представляет собой в большей степени результат аллогенных процессов, нежели аутогенного развития. В более поздней обзорной работе Оливер (Oliver, 1981) высказал предположение, что основным фактором, определяющим структуру леса и видовой состав деревьев во многих районах Северной Америки, являются сила и частота внешних возмущений.
Если в силу периодических возмущений внешней среды или вследствие характера развития самого сообщества нарушения происходят через более или менее регулярные промежутки времени, то экосистема претерпевает то, что логично было бы назвать циклической сукцессией (Уатт, 1947). Хороший пример самогенерируемой циклической сукцессии дает описанный ранее (гл. 5, разд. 4) цикл пожаров в растительности чапараля, где слаборазложившаяся подстилка образует запас горючего материала для периодических пожаров в сухой сезон.
Другой пример "волнообразной" сукцессии - высокогорные леса из бальзамической пихты на северо-востоке США (Sprugel, Bormann, 1981). Деревья здесь растут на тонком слое почвы, и по мере достижения максимальной высоты они теряют устойчивость к штормовым ветрам, которые вырывают с корнем и таким образом губят старые экземпляры. Таким образом, возникает возможность вторичной сукцессии. Как показано на рис. 8.3, склон горы покрыт рядами полос молодых, зрелых и мертвых деревьев (последние образуют на фотографии светлые полосы). Вследствие непрерывной циклической сукцессии эти полосы движутся, подобно волнам, через весь ландшафт в направлении доминирующих ветров. В любой момент времени здесь представлены все стадии сукцессии, предоставляющие множество местообитаний животным и более мелким растениям. Склон горы в целом представляет собой "циклический климакс", своего рода стационарное состояние, находящееся в равновесии со своим окружением.

Рис. 8.3. Циклическая ('волнообразная') сукцессия в лесу бальзамической пихты
Природная структура, состоящая из чередующихся полос молодого и зрелого древостоя, наводит на мысль о том, что вырубки в виде лент или пятен могут быть хорошим способом ведения лесного хозяйства на обширных пространствах, поскольку в таком случае легче происходило бы естественное возобновление (и не нужно было бы проводить дорогостоящих лесопосадок). Почвы и популяции животных также оказывались бы менее нарушенными, чем при сплошных рубках в больших лесных массивах. Кроме того, наличие разных стадий сукцессии создает множество пограничных зон, что способствует увеличению видового разнообразия диких животных.
Другой пример циклической сукцессии - цикл листовертки-почкоеда и ели, описанный в гл. 6, разд. 5. В этом случае периодические нарушения вызываются не физическими силами, а растительноядными животными, которые лишают хвои и уничтожают старые деревья, что дает возможность расти молодым деревьям.
Термин "зависимый от возмущений" часто используется для обозначения таких экосистем, которые специально адаптированы к регулярно повторяющимся возмущениям благодаря тому, что они располагают видами и процессами, способными к быстрому восстановлению (см. обзор Vogl, 1980). Для того чтобы предсказать восстановление экосистемы и управлять им после таких, например, нарушений, как открытые разработки ископаемых, необходимо в деталях знать структуру сукцессионного процесса и восстановительный потенциал конкретной экосистемы, иначе усилия по ее восстановлению могут мешать, а не способствовать естественному восстановлению (см. McIntosh, 1980).
Теоретические траектории экологической сукцессии в благоприятных и неопределенных физических условиях среды показаны на рис. 8.4. По мере развития биотического сообщества сложность организации быстро увеличивается. На увеличение содержания информации указывает возрастание биомассы и отношения В/P. Это же говорит об увеличении разнообразия и эффективности функциональных процессов, связанных с потоком энергии и циркуляцией биогенных элементов. Если физические условия среды благоприятны и вероятность катастрофических нарушений мала, то достигается высокий уровень организации и длительное время сохраняется стационарное состояние или климакс (рис. 8.4, I). Траектория, ведущая к более низкому уровню организации, характерна, напротив, для неблагоприятной среды, подверженной периодическим возмущениям, которые прерывают и возвращают вспять процесс сукцессии (рис. 8.4, II). На кривой II показаны две возвращающие сукцессию вспять петли, которые следуют после двух возмущений, обозначенных стрелками. По-видимому, этот более низкий уровень лучше адаптирован к возмущениям и более устойчив. Можно высказать рабочую гипотезу, что в общем более зрелые стадии сукцессии ведут себя стабильнее при кратковременном стрессе или стрессе, вызванном определенным фактором (как, например, засушливый год), тогда как ранние стадии устойчивее (восстанавливаются быстрее) при катастрофических стрессах, таких, как сильная буря или пожар (см. пункты 18 и 19 в табл. 8.1).

Рис. 8.4. Траектория экологической сукцессии в благоприятной среде с низкой вероятностью катастрофических возмущений (I) и в стрессовой среде с периодическими возмущениями, нарушающими и отбрасывающими назад процесс развития (II). На кривой II показаны две гипотетические петли траектории возврата сукцессии назад. Эти события происходят вслед за нарушениями, показанными стрелками; биотическое сообщество адаптируется к нарушениям, и в нем поддерживается уровень организации ниже той, которой оно бы достигло в отсутствие стресса на входе. (По Odum, 1981)
Круговороты биогенных элементов. Важной тенденцией в развитии сукцессии являются увеличение времени оборота и повышение способности к сохранению и регенерации веществ. Все это приводит к замыканию или "уплотнению" биогеохимических циклов основных биогенных элементов, таких, как азот, фосфор и кальций (табл. 8.1, пункты 7-10). Исследователи не пришли к единому мнению в оценке тех пределов, в которых сохранение биогенов представляет собой главное направление или стратегию развития экосистемы. Отчасти это противоречие обусловлено тем, что способы выявления соответствующих процессов различны. Эту проблему хорошо иллюстрирует рис. 8.5. Витусек и Рейнерс (Vitousek, Reiners, 1975) высказали предположение, что в системе, в которой биомасса накапливается на ранних стадиях сукцессии, с большей вероятностью будет идти процесс запасания биогенов. Согласно теории этих авторов, при переходе биогенов в накопленную биомассу отношение выхода к входу падает ниже единицы. В зрелой климаксной системе, где рост больше не происходит, вход и выход уравновешиваются, и это отношение вновь повышается до единицы (рис. 8.5). Вудманси (Woodmansee, 1978) отметил, однако, что даже после того, как растения перестали давать прирост живой биомассы, накопление биогенов в почве может продолжаться. К тому же Гендерсон (Henderson, 1975), Уэстмен (Westman, 1975) и Финн (Finn, неопубликованная рукопись) считают, что определение отношения выход/вход не единственный и, видимо, не лучший способ оценки поведения биогенов. Как показано на рис. 8.5, индекс возобновления (CI = отношению регенерированной части на входе к выходу, см. гл. 4, разд. 7) устойчиво возрастает по мере созревания системы. Биогены, следовательно, удерживаются более долгое время и используются повторно, при этом даже в случае сбалансированных входа и выхода необходимость притока уменьшается. Точно так же отношение запаса (S) к потерям (О), по-видимому, мало на ранних стадиях и возрастает на поздних стадиях. В общем имеются экспериментальные данные и можно также теоретически обосновать то, что в процессе развития экосистемы возрастают запас и степень возобновляемости биогенных элементов, в результате необходимый уровень поступления этих элементов на вход для поддержания единицы биомассы снижается. У нас нет оснований считать, что так же сохраняются второстепенные и токсичные вещества.
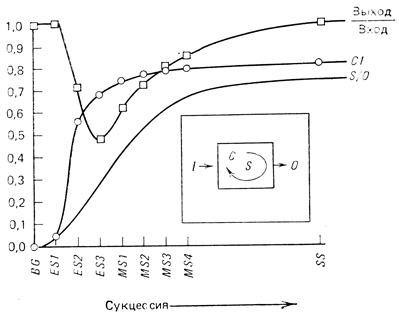
Рис. 8.5. Гипотетические тенденции изменений отношения выход/вход, индекс цикличности (CI) и отношение запас/выход (S/О) для биогенных элементов в процессе сукцессии. I - вход; О - выход; S - запас; С - возобновление; BG - обнаженная почва; ES - ранние стадии; MS - промежуточные стадии; SS - стационарное состояние. (Основано на схеме J. Т. Finn)
Райс и Пэнколи (Rice, Pancholy, 1971, 1972, 1974) представили в серии работ косвенные доказательства того, что на зрелых, или климаксных, стадиях происходит аллелохимическое ингибирование нитрификации. Таким образом, в ходе сукцессии происходит смена источника азота с нитрата на аммоний. Пионерные растения используют в первую очередь нитрат, тогда как на поздних стадиях, в частности на стадиях леса, в качестве источника азота они используют аммоний. Питер Мак-Рой (личное сообщение) наблюдал, что в зрелых стадиях сукцессии морских трав азот регенерируется в форме аммиака благодаря деятельности анаэробных микробов. Факт этот очень интересен, так как при переходе с нитрата на аммоний уменьшается количестве энергии, расходуемой на регенерацию азота (см. обсуждение круговорота азота в гл. 4), и возрастает, следовательно, эффективность использования энергии. Вопрос, однако, остается открытым, потому что Робертсон и Витусек (Robertson, Vitousek, 1981) не получили в экспериментальных данных подтверждающий сдвиг от потребления нитрата к потреблению аммония.
Не решен также вопрос о том, увеличивается или снижается в течение сукцессии интенсивность азотфиксации, микоризного симбиоза и других форм мутуализма, которые повышают эффективность регенерации биогенов (пункт 17 в табл. 8.1). Эти формы мутуализма проявляются и широко распространены на всех стадиях сукцессии, но до сих пор было предпринято мало попыток количественно оценить их деятельность на разных сукцессионных стадиях. По-видимому, мутуалистические отношения, способствующие сохранению биогенов, в большей степени подвержены влиянию "текущих запросов" (бедность биогенами), чем процессов развития экосистемы на таковых.
Замещение видов. Более или менее непрерывная смена видов во времени - характерная черта большинства сукцессионных серий. Смена видового состава растительности была названа Эглером (Egler, 1954) "флористической эстафетой". Несомненно, существует и "фаунистическая эстафета", поскольку виды животных также сменяют друг друга в сериях.
Если развитие начинается на участке, который до этого не был занят каким-либо сообществом (на обнажившихся скалах, на песке или на потоке лавы), то первичная сукцессия на ранних стадиях протекает медленно и для достижения стационарного состояния, характеризующего зрелость системы, потребуется много времени. Классический пример первичной сукцессии - дюны в штате Индиана у южной оконечности озера Мичиган, которое некогда было гораздо больше, чем сейчас. Отступая к своим современным границам, озеро оставляло все более и более молодые песчаные дюны. Поскольку субстрат песчаный, сукцессия протекает медленно, и можно наблюдать серию сообществ разного возраста: пионерные стадии у берегов и все более старые стадии па мере удаления от берега. Именно в этой "природной лаборатории сукцессии" Каулес (Cowles, 1899) провел свои основополагающие исследования растительной, а Шелфорд (Shelford, 1913) - животной сукцессии. Оба автора показали, что виды как растений, так и животных сменяют друг друга по мере увеличения возраста дюн; существующие вначале виды в старых сообществах заменяются другими, совершенно отличными от них. Олсон (Olson, 1958) повторно исследовал развитие экосистемы на этих дюнах и представил современные данные о происходящих при этом процессах и их скоростях. Из-за вторжения в эти области тяжелой промышленности специалисты по охране природы, которые пытаются сохранить "дюны Индианы", оказались в затруднительном положении. Однако, к счастью, некоторая часть этих дюн входит теперь в состав национальных парков и парков штата. Общественность должна поддержать их усилия по защите дюн, поскольку этот район не только бесценен своей природной красотой, которой могут наслаждаться жители городов, но и представляет собой естественную учебную лабораторию с поразительно наглядной экологической сукцессией.
Первые поселенцы на дюнах - это прибрежные злаки (Ammophila, Agropyron, Calamovilfa), ива, песчаная вишня, хлопковое дерево и животные, такие, как бегающие по песку длинноногие жуки-скакуны, живущие в норках пауки, кузнечики. Вслед за сообществом первых поселенцев следуют открытые сухие леса из сосны Банкса, затем черного дуба и, наконец, на самых старых дюнах - влажные леса из дуба и гикори или бука и клена. Несмотря на то что развитие сообщества начинается в очень сухом и бесплодном местообитании, оно в конечном счете приводит к влажному и холодному сомкнутому лесу, резко отличающемуся от голых дюн. Мощная богатая гумусом почва, населенная дождевыми червями и улитками, контрастирует с сухим песком, на котором она образовалась. Так сравнительно негостеприимная вначале масса песка в конце концов совершенно преобразуется под влиянием сукцессии сообщества.
На ранних стадиях сукцессия на дюнах часто приостанавливается, если ветер засыпает песком растения, и дюны приходят в движение, погребая всю растительность на своем пути. Это пример аллогенных возмущений, которые задерживают или обращают сукцессию вспять, о чем говорилось в начале данного раздела. Однако по мере продвижения дюны от берега она стабилизируется, и ее первые поселенцы, злаки и деревья, вновь восстанавливаются. Использовав современные методы радиоуглеродного датирования, Олсон (Olson, 1958) установил, что для достижения климаксного лесного сообщества на дюнах озера Мичиган требуется около 1000 лет. Это примерно в пять раз больше, чем нужно для развития зрелого леса в более гостеприимном месте, как будет видно из следующего примера.
Пример вторичной сукцессии дан в табл. 8.2, где показаны растительные сообщества и популяции птиц, сменяющие друг друга на месте заброшенных сельскохозяйственных угодий, находящихся в возвышенном предгорье на Юго-Востоке США. Первые поселенцы - это r-стратеги, однолетние растения, такие, как Digitaria, Erigeron и Ambrosia, затрачивающие большую часть своей энергии на расселение и размножение (см. табл. 6.7). Через два-три года их место занимают многолетнее разнотравье, представленное астрами и золотарником, злаки, в особенности бородач (Andropogon), и такие кустарники, как ежевика (Rubus). Если поблизости есть обильный источник семенного материала, то появляются сосны, которые вскоре образуют сомкнутый полог и затеняют ранних поселенцев. Часто вместе с соснами появляется несколько видов быстро растущих листопадных деревьев, как, например амброво и тюльпанное деревья. Поскольку все эти виды живут подолгу, стадия сосны (с редкими широколиственными деревьями) существует длительное время, но в нижнем ярусе постепенно развиваются теневыносливые дубы и гикори. Молодые сосны не могут развиваться в тени взрослых деревьев, поэтому, по мере того как старые сосны погибают от болезней, старости и ураганов, в пологе начинают доминировать дубы и гикори.
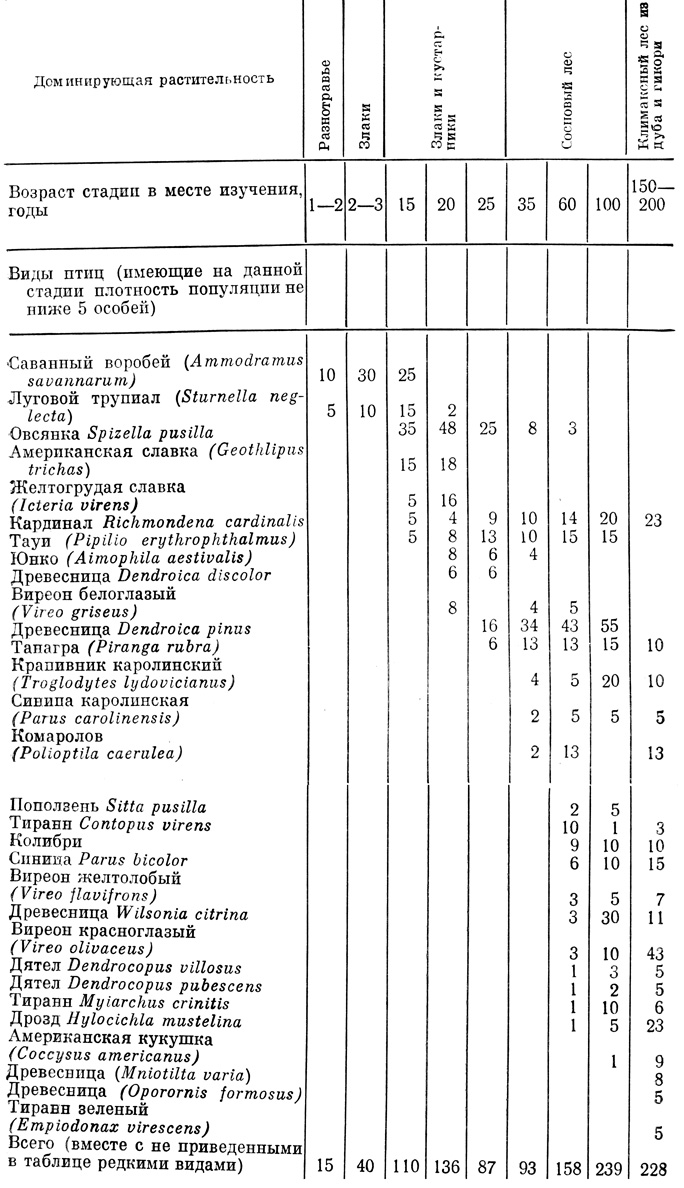
Таблица 8.2. Вторичная сукцессия в предгорной области на Юго-Востоке США*
* (По Johnston, Е. Р. Odum, 1956. Цифры показывают число занятых участков или оценку числа пар на 40 га. Главные растительные доминанты в серии, следующей за пустошами на месте полей, нарисованы над таблицей. Сукцессия была подробно описана Устингом (Oosting, 1942). Кивер (Keever, 1950) изучил некоторые взаимодействия между растениями. В таблице показана сукцессия обычно гнездящихся птиц.)
В табл. 8.2 показано, что население птиц сменяется на каждой крупной сукцессионной стадии; эти изменения проявляются сильнее всего при смене доминирующих жизненных форм растений (травы - кустарники - сосны - лиственные породы). Птицы выбирают местообитания, ориентируясь больше на жизненные формы, а не на виды растений. Ни один вид растений или птиц не может процветать на протяжении всей сукцессии. Максимумы разных, видов приходятся на разные точки временной шкалы.
Животные принимают не только пассивное участие в изменении сообщества. Птицы разносят семена, необходимые для установления стадий кустарников и широколиственных деревьев, растительноядные организмы, паразиты и хищники часто контролируют последовательность видов. И, конечно же, в мелководных морских местообитаниях именно крупные животные, а не растения образуют структурную матрицу сообщества. Глемарек (Glemarec, 1979) описал вторичную сукцессию донных животных вдали от побережья Бретани (Франция). После штормов, вызывающих перераспределение осадков и нарушающих донную фауну, наступает период относительного затишья. В это время при отсутствии внешних воздействий происходит более или менее предсказуемая смена доминирующего населения. Первыми поселяются двустворчатые моллюски, питающиеся взвесью, затем такие же моллюски, но добывающие пищу в донных отложениях, и, наконец, в бентосе начинают преобладать питающиеся детритом многощетинковые черви. Таким образом, подтверждается теория, что при непрерывной сукцессии в среде увеличивается количество органического вещества.
Вторичная растительная сукцессия в степных районах так же поразительна, как и лесная, несмотря на то что в ней участвуют только травянистые растения. В 1917 году Шанц (Shantz) описал сукцессию на старых заброшенных фургонных дорогах, по которым первые поселенцы пересекали степи центральных и западных штатов; впоследствии такую же в сущности последовательность видов описывали не один раз. Хотя в разных географических областях встречаются разные виды, общий характер сукцессии всюду одинаков. Сукцессия слагается из четырех последовательных стадий: 1) стадия однолетних сорняков (2-5 лет); 2) стадия короткоживущих злаков (3-10 лет); 3) ранняя стадия многолетних злаков (10-20 лет); 4) климаксная стадия многолетних злаков (достигается за 20-40 лет). Таким образом, природе нужно от 20 до 40 лет, чтобы, начав на голом или перепаханном грунте, "построить" степной климакс; точные сроки зависят от лимитирующих воздействий таких факторов, как влажность, выпас и т. п. Несколько засушливых лет подряд или чрезмерный выпас вызывают возврат сукцессии к стадии однолетних сорняков. Насколько далеко может зайти такой возврат, зависит от силы воздействия.
В водных местообитаниях, как и в наземных, можно наблюдать сукцессию. Но, как уже подчеркивалось, процесс развития сообщества в мелководных экосистемах (прудах и небольших озерах) обычно осложняется сильным притоком веществ и энергии, который может ускорить, остановить или повернуть вспять нормальное течение развития сообщества, т. е. такое развитие, какое наблюдалось бы без этого сильного аллогенного воздействия. Сложное взаимодействие аутогенной и аллогенной сукцессии выступает более наглядно в искусственных прудах и водохранилищах. Когда при создании водохранилища затопляют тучные почвы или участок с большим количеством органического вещества (например, при затоплении лесистой местности), первой стадией развития бывает высокопродуктивная стадия "цветения", для которой характерны интенсивное разложение, высокая активность микроорганизмов, обилие биогенных элементов, низкое содержание кислорода в придонном слое воды. При этом часто наблюдается быстрый и интенсивный рост рыбы. На этой стадии рыболовы очень счастливы. Однако после того, как запасенные биогенные элементы рассеиваются, а запасы пищи оказываются израсходованными, водоем стабилизируется на некотором более низком уровне продуктивности, более высоком содержании кислорода в придонном слое и пониженном урожае рыбы. Эта стадия приносит рыболовам огорчение (см. рис. 8.6). Такая стабилизированная стадия, своего рода климакс, может существовать некоторое время, если водосбор хорошо защищен зрелой растительностью или если почвы на площади водосбора неплодородны. Однако эрозия и различные усиливаемые человеком источники эвтрофикации обычно порождают длительную серию "переходных состояний", продолжающуюся до тех пор, пока водоем не окажется заполненным осадками. Водохранилища на истощенных водосборах или на первично стерильных участках дают, конечно, обратную, картину - низкую продуктивность вначале. Непонимание основных принципов экологической сукцессии и зависимости между водосбором и водоемом привело к многочисленным неудачам и разочарованиям при попытках человека поддерживать такие искусственные экосистемы. Мелководные экосистемы могут быть "импульсно стабилизированы" на высоком уровне продуктивности в результате поступления большого количества энергии за счет колебаний уровня воды (это будет описано в разд. 3 настоящей главы).
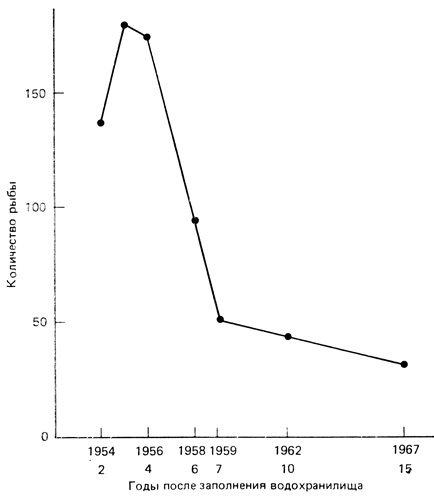
Рис. 8.6. Изменение обилия рыбы (средние по двум методам сбора) в новом русловом водохранилище в верховьях Миссури со второго по пятнадцатый годы после постройки плотины и заполнения водой образовавшегося озера Френсис-Кейз, Южная Дакота. (Gasaway, 1970)
Так как океаны, вообще говоря, находятся в стационарном состоянии и на протяжении столетий химически и биологически стабилизированы, океанографов не интересовали вопросы экологической сукцессии. Однако по мере того, как в результате загрязнений равновесие в океанах начинает нарушаться, взаимодействие аутогенных и аллогенных процессов будет, несомненно, привлекать большее внимание морских биологов. Более очевидны сукцессионные изменения в прибрежных водах, как это уже отмечалось на примере развития донных сообществ после сильных штормов, разрушающих поверхность морского дна. Маргалеф (Margalef, 1967) следующим образом подытожил свои наблюдения над сукцессионными изменениями в толще прибрежных вод:
1. Возрастают средний размер клеток и относительное обилие подвижных форм в фитопланктоне.
2. Продуктивность или скорость размножения снижаются.
3. Меняется химический состав фитопланктона, о чем можно судить, в частности, по растительным пигментам.
4. Меняется видовой состав зоопланктона: вместо пассивных фильтраторов развиваются более активные и питающиеся выборочно охотники. Это происходит вследствие сдвига от многочисленных мелких взвешенных пищевых частиц к более рассеянной пище, сконцентрированной в более крупные частицы и распределенной в более организованной (стратифицированной) среде.
5. На поздних стадиях сукцессии общий перенос энергии может быть снижен, но его эффективность, вероятно, повышается.
Джессби и Голдмен (Gassby, Goldman, 1974) использовали скорость смены видов планктона для построения диаграммы сезонной сукцессии в озерах. Виды быстро сменяют друг друга весной и намного медленнее летом. Если в июне добавить в озеро биогенные элементы, то скорость смены видов достигнет весеннего уровня и останется такой примерно в течение месяца.
Сукцессия организмов на искусственных субстратах в водной среде привлекает больше внимания, что объясняется практической важностью обрастания днищ судов и причалов усоногими ракообразными и другими сессильными морскими организмами. К тому же небольшие, используемые в нескольких повторностях образцы субстратов, такие, как стеклянные пластинки или квадраты из пластика, дерева или других материалов, широко применяются для оценки влияния загрязнений на биоту пресных и соленых вод (см. Patrick, 1954; Cairns, Dickson, 1971). Такие субстраты представляют собой своего рода микрокосм, на котором можно ожидать развития экологической сукцессии. Но это только ограниченная и упрощенная модель, поэтому экстраполировать полученные с ее помощью данные на более обширные, менее ограниченные в пространстве открытые системы со многими типами субстратов следует с осторожностью. В общем, первыми заселяют субстрат виды, у которых в том месте и в то время, где и когда появилась доступная поверхность, в воде имеются многочисленные стадии расселения. Иногда эти первые поселенцы так меняют химическую и физическую природу субстрата, что облегчается вторжение других видов, но чаще они сопротивляются вторжению других видов и существуют до тех пор, пока их не сменят более удачливые конкуренты. Как уже отмечалось при обсуждении литоральных сообществ на скалистых берегах, в процессе распределения видов в местообитаниях с определенными ограничениями или в условиях нехватки свободного пространства отрицательные взаимодействия (конкуренция, хищничество) играют большую роль, чем положительные (сосуществование, мутуализм).
Гетеротрофная сукцессия. В качестве примера гетеротрофной сукцессии можно привести данные ранних экспериментов Вудраффа (Woodruff, 1912) с сенным отваром. Если дать постоять культуральной среде, приготовленной из отвара сена, то развивается пышная культура бактерий. Если затем добавить к ней немного прудовой воды, содержащей "посевной материал" разнообразных простейших, то произойдет вполне определенная сукцессия населения простейших со сменой доминантных форм (рис. 8.7). В этой ситуации количество энергии достигает максимума вначале, а затем уменьшается. Если не добавлять свежую культуральную среду или не переводить систему в автотрофный режим, то она в конечном счете прекращает свое существование и все организмы погибают или образуют покоящиеся стадии (споры, цисты и т. п.). Такая система радикально отличается от автотрофной сукцессии, в которой поток энергии поддерживается неопределенно долго. Микрокосм в сенном отваре представляет собой хорошую модель сукцессии, происходящей в разлагающейся древесине, трупах животных, фекальных пеллетках и на вторичных стадиях очистки сточных вод. Эту сукцессию можно рассматривать также как модель сукцессии, "скатывающейся по наклонной плоскости", которую следует ассоциировать с обществом, зависящим от ископаемого горючего. Во всех этих примерах мы имеем дело с рядом переходных состояний в понижающемся энергетическом градиенте, причем стационарного состояния достичь невозможно.
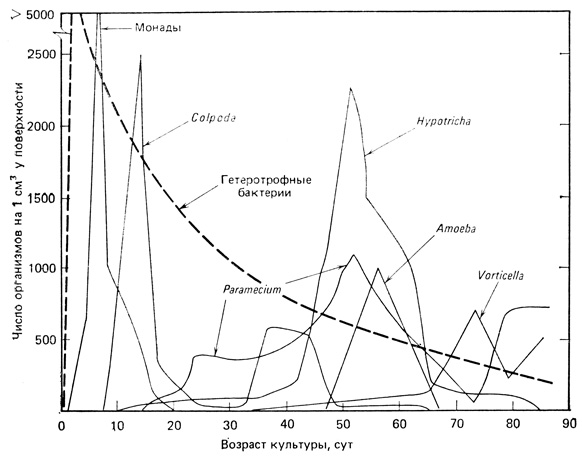
Рис. 8.7. Сукцессия в сенном отваре с последовательным доминированием разных видов. Это лабораторный пример гетеротрофной сукцессии. (По Woodruff, 1912, с добавлением гипотетической кривой для гетеротрофных бактерий)
В лабораторной модели микроэкосистемы можно скомбинировать гетеротрофную и автотрофную сукцессии. Для этого пробы из каждой системы нужно добавить в среду, обогащенную органическим веществом. Горден и др. (Gorden et al., 1969) описали сукцессию в такой системе и измерили в ней поток энергии. Вначале, когда "цветут" гетеротрофные бактерии, система становится мутной; затем, когда в результате активности бактерий в среде накапливаются биогены и ростовые вещества (в частности, витамин тиамин), нужные водорослям, система становится ярко-зеленой. Эта сукцессия служит, конечно же, хорошей моделью культурной эвтрофикации, вызываемой органическим загрязнением, таким, как неполностью очищенные городские сточные воды.
Давление отбора: количество против качества. Стадии заселения островов, описанные Мак-Артуром и Уилсоном (McArthur, Wilson, 1967), прямо соответствуют стадиям экологической сукцессии на континентах. На ранней стадии колонизации острова, когда плотность населения мала, преобладает, как и на ранних стадиях сукцессии, r-отбор, т. е. успешнее вселяются виды с высокой скоростью размножения и роста. Напротив, при равновесной плотности, характерной для поздних стадий заселения острова и экологической сукцессии, давление отбора благоприятствует K-стратегам, т. е. видам с низким потенциалом роста, но более высокой способностью к выживанию в условиях конкуренции (пункт 14 в табл. 8.1).
Можно предположить, что происходящий при сукцессии переход от количества продукции к качеству сопровождается генетическими изменениями, охватывающими всю биоту, на что указывает тенденция к увеличению размеров организмов (пункт 16 в табл. 8.1). Изменение размеров растений, по-видимому, связано с возникновением адаптации к переходу на другой источник пищи, от неорганических биогенных элементов к органическим веществам. Мелкие организмы обладают селективным преимуществом в среде, богатой биогенами и компонентами минерального питания. Это справедливо, в частности, для автотрофов, поскольку у них велико отношение поверхности к объему. Однако по мере развития экосистемы неорганические биогенные элементы оказываются все более и более связанными в биомассе (т. е. становятся интрабиотическими), так что селективное преимущество переходит к более крупным организмам (либо к более крупным особям того же вида, либо к более крупным видам, либо и к тем и к другим), способность к запасанию веществ у которых выше и жизненные циклы сложнее. Благодаря этим свойствам они адаптированы к использованию сезонных или периодических поступлений биогенов или других ресурсов.
Тенденции изменения разнообразия. Хотя оба компонента разнообразия (пункты 12 и 13 в табл. 8.1) почти всегда возрастают на ранних стадиях развития экосистемы, все же пик разнообразия в одних случаях приходится примерно на середину серии, в других - ближе к концу. Тенденции изменения разнообразия во время сукцессии неодинаковы в различных трофических или систематических группах. Ранее мы уже кратко описали основные стадии сукцессионной серии на залежных полях в предгорьях в Джорджии (табл. 8.2). Никольсон и Монк (Nicholson, Monk, 1974) определили видовое богатство и выровненность для видов растений, принадлежащих к четырем основным жизненным формам - травы, лианы, кустарники и деревья - в этих стадиях. Видовое богатство в каждом ярусе возрастает быстро, сразу после его закрепления, а затем на последующих стадиях сукцессии скорость роста этого показателя уменьшается. Выровненность сразу возрастает почти до максимума, а в дальнейшем меняется очень мало. На рис. 8.8 показаны кривые доминирования - разнообразия другой залежной сукцессии (южный Иллинойс). Разнообразие видов растений в ходе сукцессии в общем увеличивается, достигая максимума в начале стадии развития леса. Графики видового распределения в первые годы вторичной сукцессии имеют характер геометрических кривых (приближаются к прямым в полулогарифмических координатах), затем, по мере увеличения числа новых видов, постепенно становятся логнормальными. Результатом этого процесса оказывается высокий уровень выровненности (см. гл. 7, разд. 6). Последовательность кривых на рис. 8.8 очень напоминает последовательность от Арктики до тропиков, показанную на рис. 7.18.
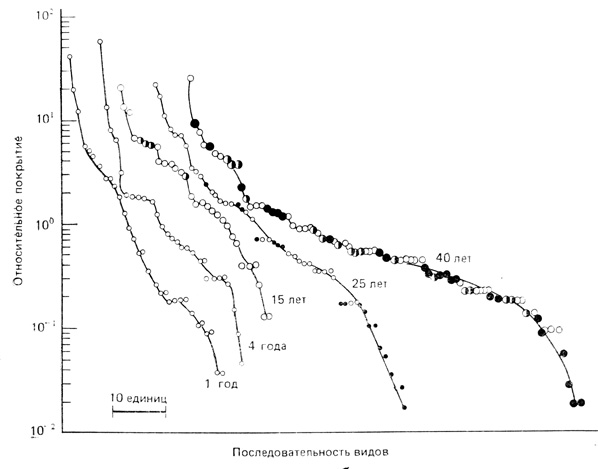
Рис. 8.8. Кривые доминирования - разнообразия для пяти разновозрастных залежных полей в южном Иллинойсе. Черные кружки - деревья, белые - травы, черные с белым - кустарники. (По Bazzaz, 1975)
Оклер и Гофф (Auclair, Goff, 1971) провели одно из наиболее полных исследований тенденций изменения разнообразия в ходе продолжительной сукцессии. Они измеряли видовое разнообразие деревьев в лесах разного возраста на западе района Великих озер. В сырых местах большее разнообразие наблюдалось в лесах среднего возраста и меньшее - в самых старых лесах. В сухих местах, напротив, разнообразие непрерывно повышалось по мере увеличения возраста леса.
Характер изменения видового разнообразия во время сукцессии - непрерывный рост или прохождение пика в промежуточной стадии - зависит от соотношения двух процессов, а именно от возрастания числа потенциальных ниш в результате увеличения биомассы, стратифицированности и других последствий биологической организации, и от противоположных эффектов увеличения размеров организмов (пункт 16 в табл. 8.1) и конкурентного исключения среди хорошо адаптированных долго живущих доминантов, которые снижают видовое разнообразие. Никому еще не удалось составить полный перечень всех видов на площади сколько-нибудь значительных размеров, не говоря уже о том, чтобы проследить за всем разнообразием видов в сукцессионной серии. Мы располагаем лишь данными для отдельных частей сообщества (деревья, птицы и т. д.). Можно ожидать, что характер изменения видового состава в значительной степени зависит от изучаемой группы и географического положения, что определяет набор видов, способных к вселению.
Как указывалось ранее (гл. 2, разд. 6), экологи единодушны в своем мнении относительно того, что изменение видового разнообразия представляет собой в большей степени косвенное следствие увеличения количества органического вещества и сложности, а не прямое следствие причинных факторов сукцессии. Достигнутый уровень разнообразия может сильно зависеть от энергетики, поскольку поддержание высокого разнообразия требует энергетических затрат и этот уровень может быть дестабилизирован (опять синдром "хорошенького понемножку").
Другие аспекты биохимического разнообразия в целом изучены далеко не столь систематически, как видовое разнообразие, тем не менее было бы логично выявить их тенденции в ходе аутогенного развития экосистемы. Например, Джеффрис (Jeffries, 1979) обнаружил, что по мере созревания и усложнения морского сообщества становится разнообразнее состав жирных кислот планктона и бентоса. Это может служить примером увеличения биохимического разнообразия.
Некоторые теоретические соображения. В начале этого раздела было установлено, что развитие экосистемы происходит в результате 1) изменений физической среды под воздействием сообщества как целого и 2) конкурентных взаимодействий и сосуществования между слагающими сообщество популяциями. Логично было бы предположить, что в многоплановое сукцессионное развитие, описанное в этом разделе, вносят вклад процессы, протекающие и на экосистемном, и на популяционном уровнях, однако" некоторые экологи считают, что такое развитие обусловлено какой-либо одной из этих причин, но не обеими. Коннелл и Слейтер (Connell, Slayter, 1977) сравнили две теории: 1) "модель облегчения", в соответствии с которой виды ранних сукцессионных стадий меняют условия существования и подготавливают, таким образом, почву для последующих вселенцев, и 2) "модель ингибирования", согласно которой первые виды противостоят вторжениям и сохраняются до тех пор, пока не окажутся вытесненными в результате конкуренции, выедания и внешних возмущений. Коннелл и Слейтер явно предпочитают вторую модель, по крайней мере для вторичной сукцессии. Сущность аргументов сторонников популяционных теорий причин сукцессии сводится к тому, что если наблюдаемые сукцессионные тенденции можно объяснить взаимодействиями на видовом уровне, то нет необходимости привлекать процессы на высших уровнях. Другие теоретики, напротив, считают, что видовая сукцессия - это только часть процесса, что самоорганизованное развитие - это свойство целых экосистем и что, следовательно, нет необходимости объяснять фундаментальные тенденции взаимодействиями составляющих популяций.
Идея о том, что экологическую сукцессию следует рассматривать с холистической точки зрения, восходит к Фридерику Е. Клементсу и его монографии 1916 г. (F. Е. Clements, 1916), озаглавленной "Растительная сукцессия" (переизданной в 1928 г. под названием "Растительная сукцессия и индикаторы"). Клементс считал, что сообщество в своем развитии повторяет последовательные стадии развития организма и что все сообщества в одной климатической области развиваются в направлении одного и тоге же климакса (идея "моноклимакса", см. следующий раздел); однако в настоящее время эти идеи потеряли свою привлекательность или оказались сильно модифицированными. Тем не менее его основной тезис - экологическая сукцессия - это процесс развития, а не только смена функционирующих изолированно видов - остается в основе одной из самых важных объединяющих теорий экологии. Маргалеф (Margalef, 1963, 1963а, 1968) и Ю. П. Одум (Е. Р. Odum, 1969) переработали и расширили фундаментальную теорию Клементса, дополнив ее такими функциональными характеристиками, как метаболизм сообщества. Главным для холистического подхода является вопрос о самоорганизации как свойстве неравновесных термодинамических систем (т. 1, с. 109). Ниже дается подборка современных утверждений, принадлежащих авторам, которые нашли по крайней мере теоретические основы для того, чтобы рассматривать сукцессию как стратегию самоорганизации: "Полагают, что процесс в целом [сукцессия] протекает таким образом, что система накапливает разнообразие и специализацию до тех пор, пока уровень неопределенности окружающей среды не обратит дальнейшее усложнение организации во вред системе". (Valiela, 1971.)
"Твердо установлено, что в стабильных условиях окружающей среды экологические системы с большим разнообразием успешно конкурируют с более простыми и замещают их. Если особый поток энергии не направлен на предотвращение сукцессии, то сложная специализированная система характеризуется большей работой выхода, направленной в русло общих усилий, и вытесняет те системы, энергия которых запасается, а не используется для полезной работы, направленной на выживание в конкуренции". (Н. Т. Odum, 1971.)
"Смена направлений приложения энергии при ее превращениях объясняет относительные эффективности видов, которые сменяют друг друга во вторичной сукцессионной серии наземных растений. Применение принципа смены способностей видов в серии позволяет предсказать самые общие свойства роста и размножения в ходе сукцессии". (Christopher Smith, 1976.)
"Ход и особенности вторичной сукцессии выводятся из теоретической динамической модели взаимообусловленных обратных связей. Эта модель [соответствует] табличной модели экологической сукцессии Одума". (Gutierrez, Fey, 1975).
"Популяции сменяются в ходе сукцессии, но продуктивность системы можно измерять на протяжении всего процесса. Сукцессия - это не просто последовательность разных систем, а единая система, которая меняет во времени переходящие виды и популяции". (O'Neil, Reichle, 1977).
"Предполагается, что самоорганизующиеся диссипативные системы развиваются во времени так, чтобы оптимизировать свою независимость (сложность биомассы и потоков в сетях). Это предположение подтверждается тенденциями развития экосистем". (Ulanowicz, 1980).
Противоположная концепция состоит в том, что экологическая сукцессия не имеет оптимизирующей стратегии и происходит в результате борьбы особей и видов за пространство. Эту концепцию высказывает в своих работах Глисон (Н. A. Gleason), в частности в статье 1926 г., озаглавленной "Индивидуалистическая концепция растительной сукцессии". Работы Глисона, рассмотренные недавно Мак-Интошем (McIntosh, 1975), служат отправной точкой построения новых популяционных теорий сукцессии. Эти теории характеризуются новыми подходами к эволюционной биологии и оценками тех влияний, которые оказывают друг на друга консументы и продуценты, и реакций на такие воздействия. Теории сукцессии, основывающиеся на свойствах организмов, а не на эмерджентных свойствах экосистем, разработаны в обзорах Друри и Нисбета (Drury, Nisbet, 1973) и Хорна (Horn, 1974, 1975). Главная предпосылка, согласно Пикетту (Pickett, 1976), состоит в том, что меняющееся в зависимости от внешних возмущений и физических градиентов положение вида в сукцессионной последовательности определяется его эволюционной стратегией (дарвиновский отбор, конкурентное исключение и т. д.) и характеристиками жизненного цикла. С тех пор как холистическая теория Клементса была признана эволюционной теорией сообщества, популяции и экосистемы, экологи не смогли далее оставаться в стороне от этой проблемы, о чем свидетельствуют их работы. Эту позицию занимал в общем покойный Роберт Уиттэкер (Whittaker, 1957, 1975; Whittaker, Woodwell, 1971). Глессер (Glasser, 1982) в недавнем обзоре также отметил, что, хотя в фазе ранней колонизации закрепляются случайные организмы, которым повезло, позднейшие стадии намного более организованны и упорядоченны.
Раньше или позже любая теория проверяется в практическом мире прикладной науки, например в лесном хозяйстве. Лесоводы, вообще говоря, считают, что лесная сукцессия направлена и предсказуема. Для того чтобы оценить будущие запасы древесины, они часто строят модели, в которых сочетаются тенденции природной сукцессии с нарушениями и хозяйственной деятельностью, меняющими природный ход процесса. В предгорных районах Джорджии, например, естественная лесная сукцессия направлена от сосен к широколиственным деревьям. Поскольку сосны в настоящее время представляют большую промышленную ценность, чем широколиственные породы, были предприняты попытки остановить сукцессию на стадии сосен, чтобы эта стадия сохранялась и восстанавливалась, особенно в районах с развитыми заготовками древесины. Джонсон и Шарп (Johnson, Sharpe, 1976) пишут, что, несмотря на усилия по освоению лесов и сохранению сосны, с 1961 по 1972 г. увеличилась площадь произрастания широколиственных деревьев (как амбрового и тюльпанного деревьев на ранних стадиях, так и дуба и гикори на поздних). Основываясь на модели, построенной на 30-летнюю перспективу, эти авторы считают, что площадь, занятая широколиственными породами, будет увеличиваться и дальше, хотя скорость такого увеличения будет меньше, чем в ходе естественной сукцессии. Важное значение в этой прогностической модели придается таким факторам, как урбанизация и борьба с пожарами, которые дают широколиственным деревьям преимущество перед соснами. Джонсон и Шарп пришли к выводу, что хотя состав предгорного леса и находится под сильным влиянием хозяйственной деятельности человека, будущие изменения будут следовать тенденциям природной сукцессии.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'