Объяснения и примеры
Термин "местообитание" широко используется не только в экологии, но и в других областях науки. Так, местообитание водных насекомых: клопа-гладыша (Notonecta) и плавта (Corixa), показанных на рис. 7.12, - это мелководные, покрытые растительностью пространства (литоральная зона) прудов и озер, куда можно пойти, чтобы собрать этих замечательных водных клопов. Эти виды занимают, однако, очень разные трофические ниши. Гладыш - активный хищник, а плавт питается преимущественно разлагающейся растительностью. Другой пример разделения ниш, основанного на способах питания, показан на рис. 7.4. В экологической литературе можно найти множество подобных примеров сосуществования видов, использующих разные источники энергии.

Рис. 7.12. Два водяных клопа (Hemiptera) - Notonecta (А) и Corixa (Б), которые живут в одном и том же местообитании, но из-за различий в питании занимают разные трофические ниши
Местообитание может также обозначать место, занимаемое целым сообществом. Например, местообитание шалфейного сообщества песчаной степи - это гряды холмов с песчаной почвой, тянущиеся вдоль северных берегов рек на юге Великих равнин в США. В данном случае местообитание слагается преимущественно из физических, или абиотических, комплексов, тогда как местообитание упомянутых выше водных клопов включает живые и неживые объекты. Таким образом, местообитание одного организма или группы организмов (популяции) включает помимо абиотической среды также и другие организмы. Для описания местообитания сообщества достаточно только последнего. Во избежание недоразумений важно различать эти два возможных применения термина "местообитание".
Экологическая ниша - понятие более новое и оно не имеет столь широкого применения вне экологии. Расплывчатые, хотя и полезные термины, такие, как ниша, трудно определить и выразить количественно; в данном случае лучше всего рассматривать историю возникновения понятия. Джозеф Гриннелл (Joseph Grinnell, 1917, 1928) пользовался словом ниша "для обозначения основной единицы распределения, в пределах которой данный вид удерживают его структурные и инстинктивные ограничения...; на одной и той же территории не может быть двух видов, которые долго занимали бы совершенно идентичную экологическую нишу". (По случайному совпадению, последнее утверждение предшествовало экспериментам Гаузе, продемонстрировавшим принцип конкурентного исключения; см. рис. 7.2.) Таким образом, Гриннелл трактовал нишу преимущественно как микроместообитание или то, что мы назвали бы теперь пространственной нишей. Чарлз Элтон (Ch. Elton, 1927, и последующие публикации) в Англии был одним из первых, кто придал термину "ниша" смысл "функционального статуса организма в сообществе". Поскольку Элтон имеет очень большое влияние на экологическое мышление, представление о том, что ниша не есть синоним местообитания, получило широкое распространение, а поскольку он уделял основное внимание энергетическим отношениям, его вариант этого понятия можно обозначить как трофическую нишу.
В 1957 г. Хатчинсон показал, что нишу можно рассматривать как многомерное пространство или гиперобъем, в пределах которого условия среды позволяют существовать особи или виду неопределенно долго. Нишу Хатчинсона, которую мы будем называть многомерной, или гиперпространственной, нишей, можно описать с помощью количественных характеристик и оперировать с ней математически. Например, двумерные климаграммы (рис. 5.15), изображающие климатическую нишу одного вида птиц и одного вида плодовой мухи, можно расширить путем введения ряда новых координат, учитывающих другие параметры окружающей среды. Хатчинсон (Hutchinson, 1965) различал также фундаментальную нишу - наибольший "абстрактно заселенный гиперобъем", когда вид не ограничен конкуренцией с другими видами, и реализованную нишу - меньший гиперобъем, занятый в условиях биотических ограничений. Эти два понятия двумерных ниш иллюстрируются на рис. 7.13, А и В.
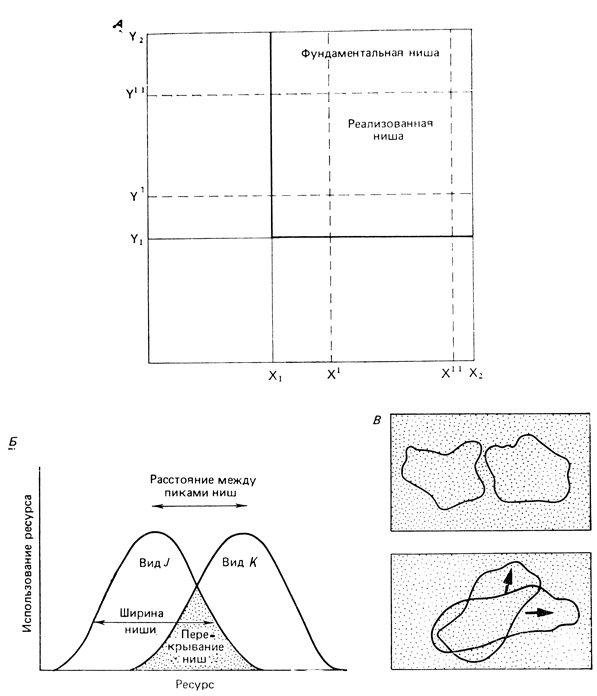
Рис. 7.13. Схематическое представление концепций ниши. А и В - ниша как гиперобъем. На верхней диаграмме В два вида занимают не перекрывающиеся ниши, на нижней диаграмме ниши перекрываются настолько сильно, что возникает жестокая конкуренция, приводящая к дивергенции (показано стрелками). Б. Распределение активности двух видов вдоль оси, имеющей размерность ресурса, иллюстрирует концепцию ширины ниши и перекрывания ниш
Вандермеер (Vandermeer, 1972) несколько расширил концепцию реализованной ниши Хатчинсона, включив в нее допущение о том, что если в данном местообитании имеется 0, 1, 2, 3, ... n взаимодействующих видов, то их реализованные ниши будут различными. В отсутствие потенциальных конкурентов вид занимает оптимальную (или фундаментальную, по Хатчинсону) нишу, но при наличии других сходных видов следует говорить не об одной реализованной нише, а о целом ряде частных ниш, число которых зависит от числа потенциальных конкурентов. Итак, Вандермеер рекомендует пользоваться термином частная ниша вместо термина "реализованная ниша". Простая аналогия из повседневной жизни, вероятно, может помочь нам разобраться в этих перекрывающихся и иногда противоречивых вариантах применения в экологии термина "ниша". Если в нашем человеческом обществе мы хотим с кем-то познакомиться, то для начала нам необходимо узнать адрес этого человека, т. е. место, где его можно найти. "Адрес" здесь соответствует местообитанию организма. Однако, чтобы узнать человека как следует, желательно знать о роде его занятий, его интересы, знакомых и вообще роль, которую он играет в жизни общества. В данном случае профессия будет аналогом ниши.
Сходная ситуация наблюдается при изучении организмов: знание местообитания - это только начало. Для определения статуса организма в природном сообществе мы должны располагать некоторыми сведениями о его активности, в частности о питании, источниках энергии и путях ее распределения; нужно, кроме того, знать соответствующие популяционные характеристики, такие, как внутренняя скорость роста, приспособленность и т. д., и, наконец, нужно знать о влиянии данного организма на другие организмы, с которыми он взаимодействует, и о той степени, в какой он воздействует или может воздействовать на важные события в экосистеме.
Мак-Артур (MacArthur, 1968) отметил, что экологический термин "ниша" и генетический термин "фенотип" - понятия аналогичные; оба они связаны с неопределенным числом признаков, имеют некоторые или даже все общие переменные и оба весьма полезны при установлении различий между особями и видами. Таким образом, ниши сходных видов, объединенных одним общим местообитанием, можно точно сравнить, если проводить сравнение лишь по нескольким операционально значимым переменным. Мак-Артур сравнивает ниши четырех видов американских лесных певунов (Parulidae). Все они размножаются в одном макроместообитании - еловом лесу, но собирают корм и гнездятся на разных частях елей. Для этой ситуации Мак-Артур построил математическую модель, представляющую собой систему уравнений конкуренции в матричной форме. Коэффициенты конкуренции в модели вычислялись для взаимодействий каждого вида с каждым из трех других. В одной из пар видов наблюдалась особенно жестокая конкуренция, поэтому весьма вероятно, что в отсутствие любого из этих двух видов другой займет вакантное пространство ниши. Общая тенденция уменьшения ниши при конкуренции между видами иллюстрируется на рис. 7.3.
Термин гильдия в современной экологической литературе часто используется в применении к группам видов, таким, как изученные Мак-Артуром лесные певуны, роль которых в сообществе одинакова или сравнима (см. Root, 1967, который первым указал на возможность такого использования термина). Можно привести и другие примеры гильдий - это осы, паразитирующие на популяциях питающихся нектаром насекомых, улитки, живущие в листовой подстилке леса, и лианы, проникающие в полог тропического леса. Гильдия - удобная единица для изучения взаимодействий между видами, однако при анализе сообщества ее можно рассматривать как функциональную единицу, в результате чего отпадает необходимость рассматривать все до единого виды по отдельности.
Разделение микроместообитаний или пространственных ниш внутри гильдий широко распространено, являясь почти правилом. Два примера показаны на рис. 7.14. У шести из семи видов лесных слизней более четко разделяются ниши в областях питания, тогда как у пяти видов бокоплавов наблюдается значительное перекрывание ниш вдоль градиента солености.
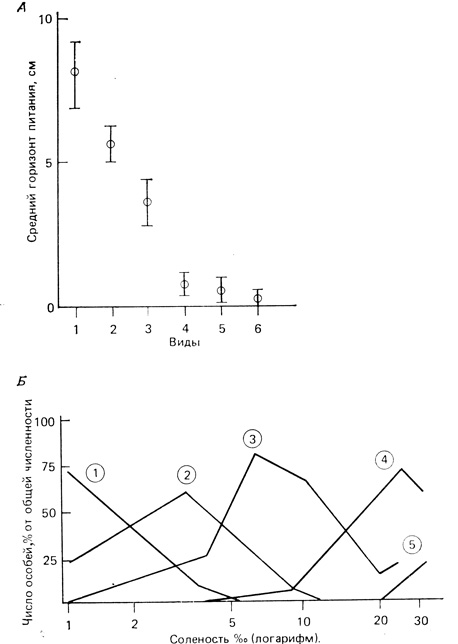
Рис. 7.14. А. Разделение ниш по высоте обитания у лесных слизней. (По Junnings, Barkham, 1979.) Б. Расположение местообитаний бокоплавов вдоль градиента солености. (По Fenchel, Raiding, 1979)
В качестве показателей при сравнении ниш крупных растений и животных часто пользуются результатами промеров морфологических признаков. Например, Ван Вален (Van Valen, 1965) нашел, что показателем "ширины ниши" могут служить длина и толщина клюва у птиц (клюв, конечно, отражает вид потребляемой пищи). Оказалось, что коэффициенты вариации толщины клювов у островных популяций шести видов птиц больше, чем у материковых; это соответствует большей ширине ниш (большему разнообразию занимаемых местообитаний и типов пищи) на островах, где меньше конкурирующих видов. Конкуренция внутри вида часто сильно уменьшается, если разные стадии жизненного цикла занимают разные ниши. Головастики, например, - растительноядные, а взрослые лягушки, обитающие в том же самом пруду, - хищники. Разделение ниш можно наблюдать даже между разными полами. У дятлов рода Picoides (старое название - Dendrocopus) самки и самцы различаются по величине клюва и пищевому поведению (Ligon, 1968). У ястребов, как и у многих насекомых, особи разного пола заметно различаются по размерам тела и, следовательно, по характеристикам пищевых ниш.
Можно ожидать, что введение в природные экосистемы как биогенных элементов, так и токсичных химических веществ в большей степени изменяет соотношение ниш тех видов, которые наиболее подвержены этому воздействию. Так, Бакелар и Одум (Bakelaar, Odum, 1978) при изучении влияния производимого в промышленных масштабах удобрения, содержащего N, Р и K, на растительность залежи обнаружили, что в результате применения этого удобрения ширина ниши и перекрывание ниш не изменились у большинства видов, но у главного доминанта - золотарника (один из видов Solidago) ширина ниши существенно увеличилась, численность его возросла за счет растения-кодоминанта (одного из видов Aster).
В следующем разделе мы рассмотрим, в какой степени распределение видов по обилию внутри трофических уровней, таксономических групп (птицы, насекомые и т. д.) и сообществ в целом позволяет нам лучше понять природу взаимодействия ниш.
Виды, занимающие одну и ту же или похожие ниши в разных географических областях, называются экологически эквивалентными, причем в соприкасающихся областях эти виды обычно близкородственны таксономически, тогда как в несоприкасающихся областях они часто не имеют родственных связей. В разных флористических и фаунистических областях сообщества сильно различаются по видовому составу, но во всех тех местах, где независимо от географического положения физические условия одинаковы, развиваются сходные экосистемы. Эквивалентные функциональные ниши оказываются занятыми теми биологическими группами, которые имеются в фауне и флоре данной области. Так, экосистемы типа злаковников развиваются повсюду, где имеются соответствующие климатические условия, но виды злаков и травоядных могут быть совершенно различными, особенно если эти области разделены мощными барьерами. Крупные кенгуру Австралии экологически эквивалентны бизонам и антилопам Северной Америки (на обоих континентах этих животных теперь в значительной степени замещают коровы и овцы).
Табл. 7.3 и 7.4 иллюстрируют два примера экологической эквивалентности в местообитаниях морского берега и степи. Брюхоногие моллюски, сменяющие друг друга по высоте берега на побережье Северной и Южной Америки, все принадлежат к одному роду, но птицы в степях Канзаса и Чили относятся к разным семействам.

Таблица 7.3. Экологические эквиваленты в трех главных нишах четырех зон побережья Северной и Центральной Америки
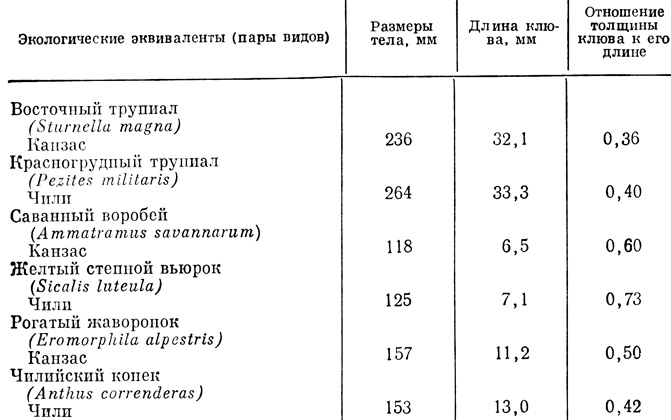
Таблица 7.4. Экологические эквиваленты среди видов степных птиц в Канзасе и Чили*
* (Три вида из каждого региона различаются своими нишами питания, на что указывают различия величины тела и размеров клюва, но каждая пара эквивалентов очень сходна морфологически, что указывает на большое сходство ниш. Трупиалы таксономически близко родственны, вторая пара родственна только на уровне семейства, виды третьей пары принадлежат разным семействам (По Cody, 1974.))
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'