Объяснения и примеры
Объективный подход к вопросу о паразитизме и хищничестве (как и к вопросу о других отрицательных взаимодействиях) найти нелегко. Все мы испытываем естественное отвращение к паразитам, будь то бактерии или ленточные глисты. Люди настроены и против всех хищников, хотя человек - самый типичный хищник и основной виновник природных эпидемий. К тому же обвинительный приговор хищнику часто выносится наобум, без выяснения того, действительно ли он вредит интересам человека. "Волк хорош только мертвым" - эта широко распространенная точка зрения, как мы увидим далее, оказывается весьма далекой от истины.
Популяционный подход наиболее объективен при анализе хищничества, паразитизма, питания растениями и аллелопатии. Хищники, паразиты и растительноядные, конечно же, убивают или наносят вред тем особям, которыми они питаются или против которых выделяют токсичные химические вещества, они замедляют до некоторой степени скорость роста популяции или уменьшают ее численность. Но означает ли это, что без хищников или химического ингибирования популяции всегда были бы в лучшем состоянии? И только ли хищники получают выгоду от связи с организмами, за счет которых они существуют, если взглянуть на эту связь с точки зрения отдаленных результатов? Как уже отмечалось при обсуждении регуляции численности популяции (гл. 6, разд. 5; см., в частности, рис. 6.20), хищники и паразиты помогают поддерживать плотность популяций растительноядных насекомых на низком уровне, так что эти последние не уничтожают собственные источники пищи и не разрушают собственные местообитания (хотя хищники могут оказаться неэффективными и в тех случаях, когда происходит вспышка численности хозяина и он ускользает от влияния зависимых от плотности регуляторных механизмов). В гл. 3, разд. 4 мы говорили о том, что эволюция растительноядных животных и растений привела к тому, что между этими организмами возникли почти мутуалистические взаимоотношения (++). Валерио (Valerio, 1975) описал случай обоюдного "хищничества" при взаимоотношениях паразит - хозяин у пауков: молодые паучки поедают паразитов, как только те выходят из яиц пауков, это позволяет паукам избежать заражения паразитами. Более полно положительные стороны паразитизма описаны Смитом (Smith, 1968).
В качестве примера популяции, которая дает вспышку численности в ответ на понижение давления хищников, обычно приводят ситуацию с оленями на плато Кайбаб, численность которых, как впервые отметил Леополд (Leopold, 1943) на основе подсчетов Расмуссена (Rasmussen, 1941), увеличилась якобы с 4000 голов (на площади 280000 га в северной части Большого Каньона, Аризона) в 1907 г. до 100000 в 1924 г., что совпало с организованной правительством кампанией по борьбе с хищниками. Коли (Caughley, 1970) повторно проанализировал имеющиеся материалы и пришел к выводу, что, хотя численность оленей действительно сильно увеличилась, популяция истощила свою кормовую базу и затем уменьшилась, вызывают сомнения степень перенаселенности, а также утверждение, что единственной его причиной явилось уничтожение хищников. Определенную роль здесь сыграли также выпас крупного рогатого скота и пожары. Коли полагает, что вспышки численности парнокопытных есть следствие таких изменений среды и условий питания, которые позволяют популяции ускользнуть от влияния обычного механизма регуляции численности.
Ясно одно: наибольшие вспышки численности отмечаются в тех случаях, когда вид интродуцируется на новую территорию, где имеются неиспользованные ресурсы и отсутствуют отрицательные взаимодействия. "Взрыв" популяции кроликов, ввезенных в Австралию, - самый известный случай такого рода среди тысяч случаев резких колебаний численности, происходящих при вселении вида с высоким биотическим потенциалом в новые места обитания. Интересное следствие попытки контроля размножения кроликов путем интродукции болезнетворных организмов предоставило нам данные, свидетельствующие о наличии группового отбора в системе паразит - хозяин (гл. 8, разд. 6).
Наиболее важное обобщение состоит в том, что отрицательные взаимодействия со временем ослабевают, если стабильность и пространственное разнообразие экосистемы достаточны для установления взаимных адаптаций. В популяциях паразитов и хозяев или хищников и жертв, интродуцируемых в экспериментальные микроэкосистемы, обычно наблюдаются резкие колебания численности с определенной вероятностью вымирания. В модельных системах хищник - жертва, описываемых уравнениями Лотки - Вольтерра, происходят постоянные незатухающие колебания, если в уравнения не введены дополнительные члены второго порядка, характеризующие действие факторов самоограничения численности, которые могут вызвать затухание колебаний. Пиментел и Стоун (Pimentel, Stone, 1968) экспериментально показали, что такие члены второго порядка могут отражать взаимные адаптации.
Как видно на рис. 7.5, сильные колебания численности происходят в том случае, когда хозяина (комнатную муху) и паразита (осу) впервые вместе помещают в замкнутую культивационную систему. Если же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, то оказалось, что у них посредством отбора выработался экологический гомеостаз, при котором каждая из популяций была "подавлена" другой в такой степени, что оказалось возможным их сосуществование при значительно более стабильном равновесии.
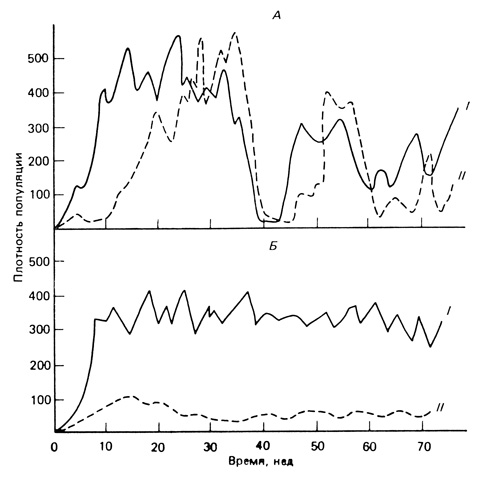
Рис. 7.5. Развитие гомеостаза в системе паразит - хозяин (хозяин - комнатная муха Musca domestica (I), паразит - оса Nasonia vitropennis (II) содержались в многокамерном садке, состоявшем из 30 пластиковых камер, соединенных друг с другом трубочками, которые замедляли распространение паразита. А. В популяциях, объединенных недавно (дикие особи впервые посажены вместе), наблюдаются сильные колебания плотности, причем эти колебания в популяции хозяина (мухи) происходят с некоторым опережением. Б. Популяции, которые получены из колоний, в которых оба вида сосуществовали в течение двух лет; отмечается более устойчивое равновесие, резкие спады численности отсутствуют. О возникновении у хозяина адаптивной устойчивости свидетельствуют сильное снижение рождаемости у паразита (46 потомков на самку вместо 133 в начале опыта) и уменьшение плотности его популяции. Этот эксперимент показывает регуляторную и стабилизирующую функции генетического механизма обратной связи в системе популяций. (А - из Pimentel, Stone, 1968; Б - по данным для двух экспериментальных популяций, приведенным на рис. 2 и 3 их работы; плотность выражена числом особей на 1 камеру в 30-камерном садке)
Однако в реальном мире обстоятельства могут складываться таким образом, что взаимная адаптация в новых ассоциациях не возникает. Всегда есть опасность, что отрицательная реакция может стать необратимой и хозяин будет элиминирован. История с заболеванием каштана в Америке служит примером такого взаимодействия популяции, которое с равной вероятностью может закончиться и адаптацией, и вымиранием.
Американский каштан раньше был важным компонентом лесов в районе Аппалачских гор на востоке Северной Америки, составляя часто до 40% биомассы леса. У него были свои паразиты, хищники и болезни. Родственный ему вид каштана, произрастающий в Китае, также имел своих паразитов, в том числе паразитический гриб Endothia parasitica, поражающий кору на стволе. В 1904 г. этот гриб был случайно завезен в США. Американский каштан оказался неустойчивым к этому новому паразиту, так что к 1952 г. все крупные деревья погибли, их мрачные серые стволы стали характерной чертой аппалачских лесов (рис. 7.6). Каштан продолжает давать корневые побеги, и эти побеги, прежде чем их снова убьет гриб, часто плодоносят, но никто не может сказать, каким будет окончательный исход этой борьбы - вымирание или адаптация. Таким образом, популяция каштана, по крайней мере на время, утратила свою былую доминирующую роль в лесной экосистеме. Есть, однако, некоторая надежда на то, что недавно обнаруженное вирусное заболевание гриба можно будет использовать для борьбы с болезнью каштанов (см. Anagnostakis, 1982). (Реакция леса на изъятие главного доминанта показана в табл. 7.7.)

Рис. 7.6. Последствие заболевания каштанов в Южных Аппалачах (Джорджия). Пример того, сколь роковым может оказаться воздействие паразитического организма (гриба), завезенного из Старого Света, на нового, инфицированного им хозяина (американский каштан)
Приведенные выше примеры не подбирались специально, чтобы подтвердить определенную точку зрения. Тот, кто интересуется этим вопросом, легко может обнаружить множество подобных примеров, проведя совсем немного времени в библиотеке. Ему станет ясно, что: 1) при длительном контакте паразитов и хищников с их хозяевами или жертвой влияние на последних становится умеренным, нейтральным или даже благоприятным (конечно, при рассмотрении длительных промежутков времени) и 2) наиболее сильное повреждающее действие оказывают новые паразиты и хищники. Действительно, если составить список болезней, паразитов и насекомых-вредителей, причиняющих наибольший ущерб сельскому и лесному хозяйству, то окажется, что многие из них недавно занесены в новые для них районы, как, например, гриб Endotia parasitica, паразитирующий на каштане, или недавно приобрели новых хозяев или жертв. Европейская хлопковая совка, непарный шелкопряд, японский жук, средиземноморская плодовая мушка - это лишь немногие из интродуцированных насекомых-вредителей, принадлежащих к этой категории. Тот же самый принцип применим во многом и к болезням человека; наиболее опасны новые, завезенные впервые возбудители заболеваний.
В общем то, что мы называем "принципом внезапного усиления патогенности", можно сформулировать следующим образом: эпидемии, эпизоотии или эпифитотии часто вызываются 1) внезапным или быстрым вселением организма, обладающего потенциально высокой скоростью собственного роста, в экосистему, в которой механизмы регуляции численности для этого нового вида или отсутствуют или малоэффективны; 2) резкими или очень сильными изменениями окружающей среды, приводящими к уменьшению энергии, необходимой для регуляции по принципу обратной связи, или каким-либо иным образом нарушающими способность системы к саморегуляции. Урок из всего этого таков: не следует допускать интродукцию новых потенциальных вредителей и избегать по возможности стресса в экосистемах, возникновению которого способствуют ядохимикаты, уничтожающие как вредные, так и полезные организмы.
Хотя с экологической точки зрения хищничество и паразитизм очень сходны, между их крайними формами - крупным хищником и мелким внутренним паразитом - имеются весьма существенные различия, касающиеся не только размеров. Паразитические или патогенные организмы обычно обладают более высоким биотическим потенциалом, чем хищники. Их строение, обмен, выбор хозяина и жизненные циклы часто более специализированы, что связано со своеобразием их среды обитания и проблемой распространения от одного хозяина к другому.
Особенно интересны организмы, занимающие промежуточное положение между хищниками и паразитами, например паразитические насекомые, которые нередко, подобно хищникам, пожирают свою добычу целиком, но вместе с тем напоминают паразитов своим высоким биотическим потенциалом и высокой специфичностью к хозяину. Энтомологи научились искусственно разводить некоторых паразитических насекомых и использовать их для борьбы с вредными насекомыми. Попытки использовать с той же целью крупных неспециализированных хищников в общем оказались неудачными. Например, мангусты, которых завезли на острова Карибского моря для уничтожения крыс на полях сахарного тростника, значительно сильнее понизили численность гнездящихся на земле птиц, чем численность крыс. Такой способ борьбы с вредителями может быть успешным при условии, что хищник мал, специализирован в выборе пищи и обладает высоким биотическим потенциалом. Хороший пример такого рода приводят в своей работе Хаффейкер и Кеннетт (Huffaker, Kennett, 1956). Эти авторы описали мелкого хищного клеща, который эффективно контролирует численность растительноядного клеща на посадках земляники в Калифорнии. Растительноядные насекомые иногда успешно используются в борьбе с сорняками (Huffaker, 1957, 1959).
Человек, к сожалению, еще очень медленно учится быть, по выражению Слободкина (Slobodkin, 1962), "расчетливым хищником", т. е. не истреблять свою жертву полностью при эксплуатации ее как ресурса. Проблема оптимального изъятия из эксплуатируемой популяции обсуждалась Бивертоном и Холтом (Beverton, Holt, 1957), Риккером (Ricker, 1958), Меншуткиным (1962), Слободкином (Slobodkin, 1962), Силлименом (Silliman, 1969), Вагнером (Wagner, 1969), Мак-Калохом (McCullought, 1979) и многими другими. Теоретически, если кривая сигмоидного роста симметрична, как в логистической модели, то скорость роста dN/dt будет максимальной при плотности, равной K/2 (при половине величины плотности насыщения). Эта точка перегиба сигмоидной кривой была обозначена как уровень поддерживающей емкости I на рис. 3.19. Однако вершина "горбатой" или параболической кривой скорости роста вследствие скошенности вправо (из-за временных задержек) часто может и не находиться точно посередине между 0 и К, поэтому плотность, при которой обеспечивается максимальный выход живой продукции, часто составляет несколько меньше половины равновесной плотности неэксплуатируемой популяции.
К анализу проблемы можно подойти экспериментально, создавая опытные популяции в микроэкосистемах. Одна из таких экспериментальных моделей представлена на рис. 7.7. Аквариумную рыбку гуппи (Lebistes reticulatus) использовали для имитации популяции промысловой рыбы, облавливаемой человеком. Можно видеть, что максимальный урожай получали в том случае, когда в каждый репродуктивный цикл изымалась одна треть популяции, при этом равновесная плотность оказывалась несколько меньшей, чем половина плотности необлавливавшейся популяции. В пределах данного эксперимента это соотношение оставалось независимым от несущей емкости системы, которая поддерживалась на трех разных уровнях путем изменения количества корма.
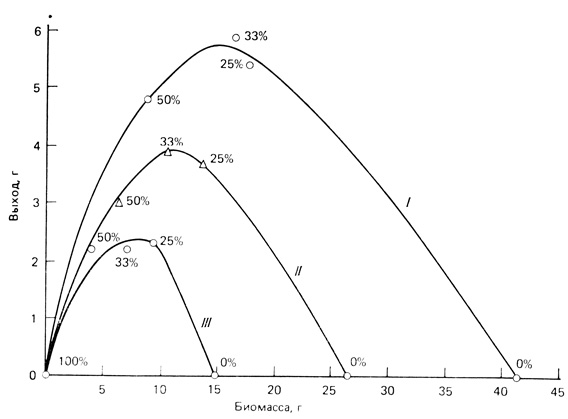
Рис. 7.7. Биомасса и выход продукции в экспериментальных популяциях гуппи, которые содержались при трех режимах питания и разных уровнях эксплуатации (в процентах изъятия за репродуктивный период; уровень доступной пищи в опыте II принят за единицу; в опыте I он в 1,5 раза больше, а в опыте III - в два раза меньше). Наибольший выход продукции получен в том случае, когда в каждый репродуктивный период изымали около трети популяции, а средняя биомасса снижалась менее чем на половину по сравнению с биомассой в неэксплуатируемой популяции (кривые выхода продукции скошены влево). (По Silliman, 1969)
Одновидовые модели часто слишком упрощают реальные отношения, поскольку в них не учитывается роль конкурирующих видов; между тем плотность последних может увеличиваться из-за уменьшения плотности изымаемого вида, и, кроме того, они используют пищу и другие ресурсы, необходимые для поддержания эксплуатируемого вида. Хищнику высшего порядка, например человеку (или наиболее эффективному пастбищному животному, например корове), очень легко нарушить конкурентное равновесие, что может привести к замещению эксплуатируемого вида на другой, непригодный для данного хищника или пастбищного животного. Случаи таких "сдвигов" все множатся, по мере того, как активизируется рыболовная, охотничья и т. п. деятельность человека. Это дает основание высказать следующее утверждение, которое следует рассматривать и как вызов и как предупреждение: одновидовые системы, как и монокультуры в сельском хозяйстве, неустойчивы по своей природе, поскольку в условиях стресса они сильно уязвимы для конкурентов, возбудителей болезней, паразитов, хищников и других факторов, оказывающих отрицательное влияние. Несколько хороших примеров этого общего принципа можно найти в промышленном рыболовстве.
Мерфи (Murphy, 1966, 1967) отметил уменьшение численности интенсивно облавливаемой популяции тихоокеанской сардины (Sardinops caerulea) и последующее возрастание численности близкого по экологии анчоуса (Engraulis mordox); в данном случае произошло замещение чрезмерно эксплуатируемого вида его конкурентом, не подвергавшимся эксплуатации.
В другом случае Смит (Smith, 1966) описал, как последовательная эксплуатация разных видов в оз. Мичиган в сочетании с интродукцией новых видов и эвтрофикацией привела к чередованию вспышек и падений численности промысловых рыб. Первоначально на протяжении полустолетия неизменным объектом рыболовства оставалась озерная форель, однако в результате чрезмерного облова, интродукции паразитической миноги и эвтрофикации форель была фактически уничтожена. Вслед за этим, быстро сменяя друг друга, здесь появлялись ряпушка, сиг, голавль и один экзотический вид сельдевых. Их численность в свою очередь сначала возрастала, а затем под влиянием усиленного отлова, конкуренции, хищников и паразитов резко снижалась. В 1960 г. в озеро был интродуцирован кижуч, который, к радости рыболовов-спортсменов, сильно размножился, питаясь экзотическим видом сельди Pomolobus pseudoharengus. Однако эта неожиданная удача может быть сведена на нет, если не будет налажен лучший контроль за количеством отлавливаемой рыбы и сбросом сточных вод.
Давление хищников или промысел часто влияют на размеры особей эксплуатируемой популяции. Так, отлов рыбы на уровне максимального урожая обычно приводит к уменьшению среднего размера рыб, а вырубка леса, при которой достигается максимальный выход пиломатериалов на единицу объема древесины, вызывает уменьшение величины деревьев, ухудшение качества древесины. Как мы неоднократно повторяли, нельзя одновременно добиться и максимального количества, и наилучшего качества. Брукс и Додсон (Brooks, Dodson, 1965) описали случай, когда в озерах крупные виды зоопланктеров заменялись более мелкими после вселения питающихся зоопланктоном рыб, которых ранее в этих озерах не было. В подобных случаях, когда экосистема относительно мала, один или несколько видов хищников способны контролировать как размерный, так и видовой состав всего трофического уровня.
Хорошие примеры аллелопатии можно найти в работе Муллера и сотрудников, которые изучали ингибиторы, продуцируемые ароматическими кустарниками в Калифорнийском чапарале. Эти исследователи не только выявили химическую природу и физиологическое действие ингибирующих веществ, но показали также их важность в регуляции состава и динамики сообщества (см. Muller, 1966, 1969; Muller et al., 1964, 1968). На рис. 7.8 показано, как летучие терпены, выделяемые двумя видами ароматических кустарников, ингибируют рост травянистых растений. Летучие токсины (особенно эвкалиптол и камфора) продуцируются в листьях и во время сухого сезона накапливаются в почве в таком количестве, что с наступлением сезона дождей они подавляют прорастание семян или последующий рост проростков в обширной зоне вокруг каждой группы кустов. Другие виды кустарников образуют водорастворимые антибиотические вещества различной химической природы (например, фенолы или алкалоиды), которые также дают им возможность занять доминирующее положение в сообществе. Однако периодические пожары, являющиеся неотъемлемой частью экосистемы чапараля, эффективно уничтожают источники токсинов, обезвреживают токсины, накопившиеся в почве, и тем самым дают возможность появиться здесь растениям, семена которых устойчивы к пожарам. В результате в дождливый сезон, следующий после пожара, наблюдается пышное цветение однолетников, которое происходит каждую весну до тех пор, пока кустарники опять не разрастутся и их токсическое действие не достигнет достаточной эффективности. В зрелом сообществе чапараля выживает очень немного травянистых растений. Таким образом, за счет попеременного действия пожаров и антибиотиков происходят циклические изменения видового состава растительности, что представляет собой адаптивную черту экосистемы этого типа.
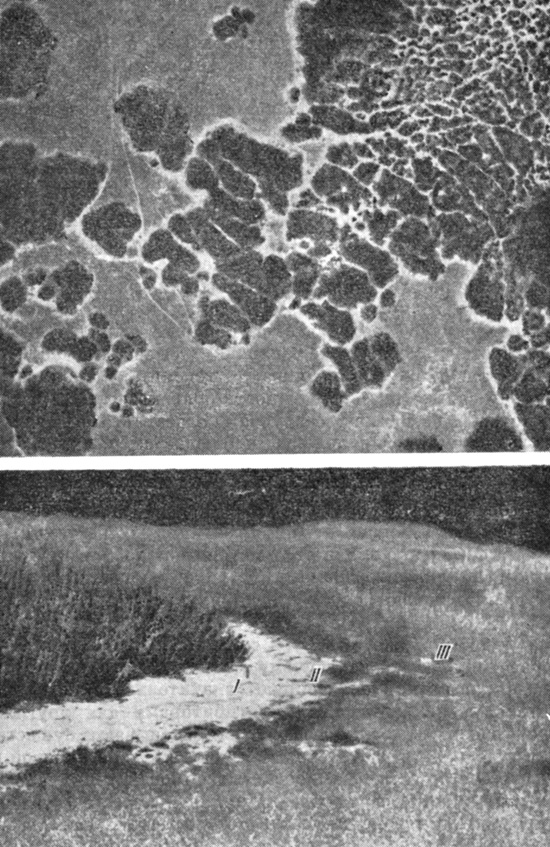
Рис. 7.8. Вверху. Аэрофотоснимок, на котором отчетливо видно вторжение ароматических кустарников Salvia leucophylla и Artemisia californica в заросли однолетних степных растений в долине Санта-Инес, Калифорния. Виден эффект биохимического ингибирования. Внизу. Куст Salvia крупным планом (слева от I), выделяющий летучие токсины и подавляющий вокруг себя рост других растений. Между I и II находится зона шириной в 2 м, лишенная всякой травянистой растительности, за исключением нескольких маленьких, чахлых проростков. Таким образом, корневая система кустарника, простирающаяся под этой зоной, не испытывает конкуренции со стороны других видов. В зоне между II и III находятся низкорослые растения, и видов здесь меньше, чем на участках справа от III, на которые не распространяется ингибирующее влияние. (Фотографии с разрешения д-ра С. Н. Muller из Калифорнийского университета, Санта-Барбара)
Уиттэкер (Whittaker, 1970) в своем обзоре растительных ингибиторов приходит к следующему заключению:
"Высшие растения синтезируют значительное число веществ, являющихся репеллентами или ингибиторами для других организмов. Аллелопатические воздействия оказывают существенное влияние на скорость растительных сукцессий и на видовой состав стабильных сообществ. Химические взаимодействия двояко сказываются на видовом разнообразии природных сообществ; сильное доминирование и интенсивные аллелопатические эффекты поддерживают низкое видовое разнообразие в одних сообществах, тогда как в других сообществах благодаря различным химическим адаптациям (вследствие того, что эти адаптации лежат в основе дифференциации ниш) поддерживается высокое видовое разнообразие".
Антибиоз, разумеется, не является достоянием только высших растений, известны многочисленные примеры его среди микроорганизмов. Иллюстрацией этого служит гриб Penicillium, образующий бактериальный ингибитор пенициллин, в настоящее время широко используемый в медицине. Химический антибиоз можно рассматривать как форму интерференционной конкуренции или, в смягченном виде, как аменсализм. Более подробно явление аллелопатии рассмотрена Райсом (Rice, 1974).
Эти интересные аспекты биологической и химической "войны" в природе вновь приводят нас к вопросу о роли отрицательных взаимодействий в экосистеме в целом. Часто значимость биотических взаимодействий зависит от положения сообщества в физическом градиенте. На рис. 7.9 представлена "модель усоногих", основанная на экспериментальном исследовании Коннелла. Литоральная зона на скалистом берегу моря предоставляет исследователю миниатюрный градиент от участков, где преобладают стрессовые физические воздействия на организм, до участков, где большее значение приобретает биологическая регуляция. В проведенных в Шотландии исследованиях Коннелл (Connell, 1961) обнаружил, что личинки двух видов усоногих ракообразных оседают в широкой полосе литоральной зоны, но доживают до взрослого состояния на более ограниченном участке литорали. Оказалось, что распространение более крупного вида (Balanus) ограничено нижней частью литоральной зоны, поскольку он не может выносить долгих периодов обсыхания. Более мелкий вид (Chthamalus) вытеснен из нижней зоны в результате конкуренции с крупным видом и более активными в нижней литорали хищниками. Таким образом, физическое стрессовое воздействие - обсыхание - оказывается основным контролирующим фактором в верхней части градиента, тогда как межвидовая конкуренция и хищничество служат главными контролирующими факторами в нижних зонах. Эту модель можно применить и к значительно более широким градиентам, таким, как Арктика - тропики или градиент от высокогорных областей до уровня моря; необходимо только иметь в виду, что все модели, хотя и в разной степени, чрезвычайно упрощают реальные отношения.
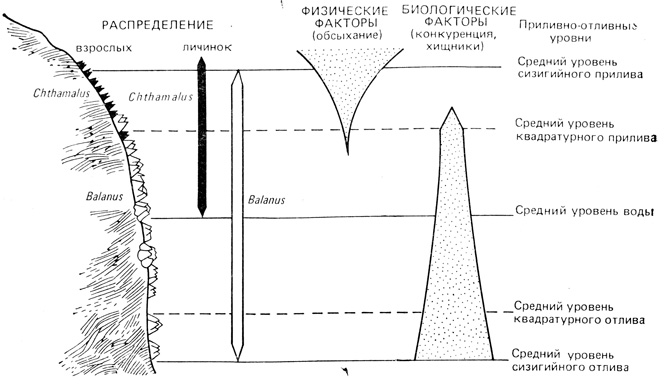
Рис. 7.9. Факторы, контролирующие распределение двух видов усоногих ракообразных в литоральном градиенте. У обоих видов взрослые особи обнаруживаются в более узкой области по сравнению с той, в которой распространяется их молодь. Распространение Balanus вверх ограничивается физическими факторами (например, обсыханием), а распространение Chthamalus вниз ограничивается биологическими факторами (конкуренция, хищничество). (По Е. Odum, 1963, из Connell, 1961)
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'