Примеры
Всем нам знакомы сезонные изменения величины популяции. Тучи москитов и комаров, леса, полные птиц, поля, заросшие амброзией, - все это бывает в свое время, в другие сезоны популяции этих организмов могут сходить практически на нет. Хотя в природе трудно найти популяции животных, микроорганизмов и травянистых растений, численность которых не изменялась бы по сезонам, наиболее выражены эти флуктуации у организмов с ограниченным периодом размножения, особенно у организмов с коротким жизненным циклом или со значительными сезонными различиями расселения в пространстве (например, у мигрирующих животных). На рис. 6.13, как уже упоминалось, показана не только J-образная кривая роста, но и сезонные и годичные флуктуации, выявленные в результате многолетних систематических исследований. Изменения такого рода, по-видимому, типичны для большинства насекомых, большинства однолетних растений и "однолетних" животных. Сезонные циклы планктонных популяций интенсивно изучались и моделировались как в пресных водах, так и в океане (один пример был описан в гл. 5, рис. 5.2).
Пример довольно нерегулярного изменения величины популяции, коррелирующего, видимо, с погодными условиями, показан на рис. 6.14. На протяжении многих лет популяция цапель в двух местностях в Великобритании оставалась относительно постоянной. Очевидно, местные условиях обеспечивали достаточно устойчивую несущую емкость среды для цапель. Однако в годы, следовавшие после лет с суровыми зимами (указаны на рис. 6.14 вверху), происходило резкое снижение плотности с последующим ее восстановлением. Синхронность изменений плотности в обеих местностях свидетельствует о том, что их причина - увеличение смертности в зимнее время. Всегда проводить исследование, не ограничиваясь только одним районом, - вот хорошее правило для полевого эколога! Популяции птиц оказались одними из наиболее хорошо изученных, поэтому именно результаты этих исследований внесли большой вклад в популяционную теорию. Обзор работ по исследованиям популяций птиц принадлежит Лэку (Lack, 1966).
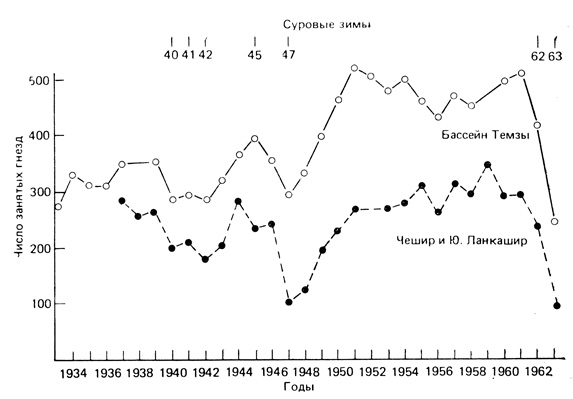
Рис. 6.14. Изменения численности цапли Ardea cinerea в двух местностях в Великобритании в период с 1933 по 1963 гг. Показана зависимость между холодными зимами и снижением численности. (Из Lack, 1966)
К наиболее известным примерам "циклических" колебаний можно отнести колебания численности у некоторых видов северных млекопитающих и птиц, у которых наблюдаются либо 9-10-, либо 3-4-летние циклы. Классическим примером 9-10-летних колебаний служит изменение численности зайца-беляка и рыси (рис. 6.15). Канадская Компания Гудзонова залива располагает начиная примерно с 1800 года ежегодными отчетами о заготовке пушнины. Если представить эти данные графически, то оказывается, что, например, у рыси пики численности популяции за весь этот длительный период повторяются каждые 9-10 лет, в среднем через 9,6 года. Пики численности часто сменяются резкими спадами, когда рысь в течение нескольких лет встречается крайне редко. У зайца-беляка наблюдаются такие же циклы, но пики его численности, как правило, на год или более предшествуют пикам рыси. Поскольку рысь в большой степени зависит от зайца, служащего ей пищей, очевидно, что цикл хищника связан с циклом жертвы. Но эти два цикла связаны не строгими причинно-следственными взаимодействиями хищник - жертва, так как циклы численности зайца отмечаются и в тех местах, где рысь не водится. Более короткие 3-4-летние циклы характерны для многих северных мышевидных грызунов (леммингов, мышей, полевок) и их хищников (особенно полярной совы и песцов). Цикл лемминга в тундре и циклы песца и полярной совы впервые на фактическом материале показал Элтон (Elton, 1942). Каждые три или четыре года на огромных пространствах тундры двух континентов численность леммингов (два вида рода Lemmus в Евразии и один в Северной Америке и еще один вид рода Dicrostonyx в Северной Америке) катастрофически возрастает, вслед за чем, часто за один сезон, резко падает. Численность песцов и сов, увеличивавшаяся по мере увеличения количества пищи, также очень скоро снижается. Совы при этом в поисках пищи могут мигрировать на юг в США (иногда достигая шт. Джорджия). Эта взрывная миграция избытка птиц происходит, по-видимому, в одном направлении; если обратный перелет и совершается, то лишь единичными особями. Таким образом, популяция сов резко уменьшается в результате миграции. Эти колебания так регулярны, что орнитологи в США могут точно предсказать нашествие полярных сов каждые три или четыре года. Поскольку эти птицы хорошо заметны и появляются повсюду вокруг городов, они привлекают к себе большое внимание: их фотографии появляются в газетах, чучела - в местных магазинах. В периоды между нашествиями ни в США, ни на юге Канады полярная сова не встречается или очень редка. Гросс (Gross, 1947) и Шелфорд (Shelford, 1943) проанализировали данные о нашествии сов и показали, что они коррелируют с периодическими уменьшениями числа леммингов - основной пищи сов.
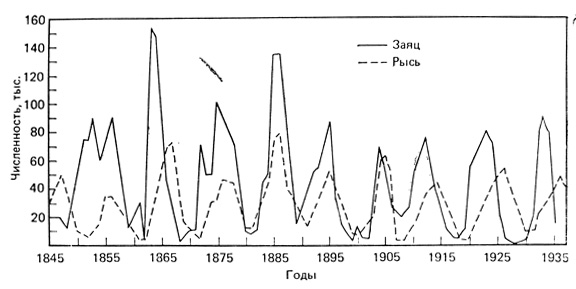
Рис. 6.15. Изменения численности рыси и зайца-беляка, регистрируемые по числу шкурок, заготовленных Компанией Гудзонова залива. Это классический пример циклических колебаний плотности популяций. (Рисунок из работы MacLulich, 1937)

Рис. 6.16. Заяц-беляк, знаменитый в истории экологии в связи с резко выраженными колебаниями численности (см. рис. 6.15). Показано животное в белом зимнем наряде. Было выявлено, что смена коричневого летнего наряда на белый зимний контролируется длительностью фотопериода. (Фотография службы охраны почв США)
В Европе (но, по-видимому, не в Северной Америке) лемминги порой достигают такой высокой плотности, что начинают мигрировать из своих перенаселенных местообитаний. Элтон (Elton, 1942) живо описал знаменитую миграцию леммингов в Норвегии. Животные проходили через деревни в таком количестве, что кошки и собаки, которые вначале нападали на них, просто переставали обращать внимание на орды грызунов. Достигнув моря, многие лемминги тонули. Движение леммингов, подобно движению сов, при взрывном возрастании их численности происходит в одном направлении. Такие впечатляющие миграции происходят не при каждом четырехлетием пике плотности, а только в годы исключительно высоких плотностей. Часто сокращение численности популяции происходит без переселения животных из тундры или с гор.
Кребс и Мейерс (Krebs, Meyers, 1974) подготовили обзор популяционных циклов у мелких млекопитающих, а Файнерти (Finerty, 1980) применил некоторые новые статистические подходы, пытаясь ответить на вопрос: "Существуют ли циклы?" Ответ гласит "да" для одних видов и ситуаций и "нет" для других.
На рис. 6.17 приведены два примера многолетней регистрации значительных колебаний численности листогрызущих насекомых в европейских лесах. Такие хорошо выраженные циклы отмечались преимущественно в северных лесах, особенно в чистых хвойных древостоях. Плотность может меняться больше чем на пять порядков (логарифмы циклов), от менее чем одной до более чем 10000 особей на 1000 м2 (верхний график на рис. 6.17), Легко представить, что 10000 потенциальных бабочек сосновой пяденицы на каждые 1000 м2, способных дать несколько поколений за сезон, более чем достаточно для появления такого числа гусениц, чтобы уничтожить всю хвою и даже погубить деревья, как это часто и случается. Циклы у этих насекомых не так регулярны, как у зайца-беляка, и повторяются примерно через 4-10 лет (в двух приведенных примерах пики численности повторялись через 7,8 и 8,8 года). У разных видов циклы не синхронны.
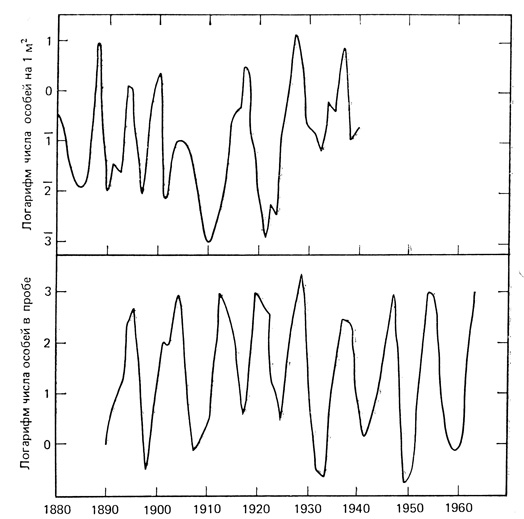
Рис. 6.17. Колебания численности двух видов бабочек, личинки которых питаются хвоей. Вверху - сосновая пяденица Bupalus в сосновом бору близ Литцлингена, Германия. В этом лесу поддерживается чистое сосновое насаждение. (Varley, 1949). Внизу - лиственничная листовертка Zeiraphera griseana в лиственничном лесу долины Энгадин, Швейцария
Хорошо известные примеры сходной периодичности - циклические вспышки численности еловой листовертки-почкоеда и коконопряда в северных районах Северной Америки. Исследования по коконопряду провел Веллингтон (Wellington, 1957, 1960), а по листовертке-почкоеду - Мак-Дональд (MacDonald, 1975) и Холлинг с сотрудниками (Ludwig et al., 1978; Clark et al., 1979; Holling, 1980). Считается, что чередование "сильных" и "слабых" физиологических рас у коконопряда (Malacosoma) имеет генетическую основу. На подъеме цикла гусеницы строят удлиненную паутину, особи активно питаются и перемещаются в ветвях на большие расстояния. В пике численности гусеницы становятся неактивными, строят более компактную паутину, едят меньше и оказываются более восприимчивыми к болезням. Таким образом, в популяции развился ограничительный контролирующий механизм, благодаря которому предотвращается повторяющаяся дефолиация деревьев и достигается длительное сосуществование насекомых и их растений-хозяев.
Можно с уверенностью сказать, что циклы листовертки-почкоеда представляют собой явления на экосистемном уровне, поскольку эволюция насекомых-листоедов, их паразитов и хищников, а также хвойных деревьев (ели и бальзамической пихты, которые часто растут в чистых насаждениях) строго сопряжена пли протекает совместно. По мере возрастания биомассы деревьев в лесу наиболее крупные и старые деревья становятся восприимчивыми к гусеницам листовертки-почкоеда, и многие из этих деревьев погибают от повторяющейся дефолиации. Отмирание и разложение древесины, фекалий и остатков насекомых возвращает в лесную почву питательные вещества. Менее чувствительные к нападению насекомых молодые деревья избавляются от затенения и быстро растут, за несколько лет заполняя полог. Тем временем паразиты и птицы, питающиеся насекомыми, совместными усилиями снижают экологическую плотность листовертки-почкоеда. Если рассматривать существование хвойных лесов на больших промежутках времени, то очевидно, что листовертка-почкоед, периодически омолаживая экосистему хвойного леса, представляет собой неотъемлемую часть этой целостной системы, а не являет собой катастрофу, как могло бы показаться всякому, кто видит только мертвые и умирающие деревья в пике цикла. Действительно, Питермен (Peterman, 1978), изучив роль короедов в полегании жердняка в горных лесах, пришел к заключению, что благодаря деятельности жуков, которые изреживают загущенный древостой, малопригодный для лесозаготовок, обитания в нем животных и отдыха, создаются более ценные для человека леса. Пожалуй, Питермен заставляет нас рассматривать этих жуков как одно из средств ведения хозяйства, а не как вредителей леса. По его мнению, в изолированных лесах следовало бы допускать периодические вспышки численности жуков. Такой взгляд, естественно, противоречит традиционным представлениям о мерах борьбы с вредителями, поскольку эти меры предусматривают контроль численности насекомых только в периоды, когда их становится слишком много и они способны полностью уничтожить деревья. Другая стратегия может заключаться просто в том, чтобы использовать в хозяйстве старые рослые деревья, прежде чем их истребят жуки или листоеды. Сейчас такая стратегия входит в практику, поскольку созданы модели, на основе которых можно предсказать время вероятной вспышки численности насекомых. Периодические бури выполняют функцию, подобную той, что выполняют насекомые: вываливая старые деревья и разрежая загущенные древостой горного леса, бури создают пятнистость в размещении молодых и более старых лесов, которые постоянно смещаются по склону (см. гл. 8, рис. 8.3).
Во всех описанных до сих пор случаях взаимодействующие популяции животных и растений длительное время эволюционировали совместно и, таким образом, выработали стратегии не только для выживания, но и для взаимной выгоды.
Пожалуй, наиболее известные примеры колебаний численности у насекомых - ее циклы у саранчовых. Сведения о нашествиях странствующей саранчи (Locusta migratoria) в Евразии восходят к далекой древности (Carpenter, 1940а). Саранча живет в пустынях и полуаридных зонах и на протяжении многих лет не совершает миграций, не пожирает посевов и не привлекает к себе внимания. Однако время от времени плотность популяции саранчи достигает чудовищных размеров. Под влиянием скученности насекомые претерпевают морфологические изменения* (например, у них развиваются более длинные крылья) и начинают мигрировать в земледельческие районы, съедая все на своем пути. Уваров (Uvarov, 1957) отметил, что такая практика возделывания земель, как севообороты и перевыпас скота, не уменьшают, а увеличивают вероятность вспышки, потому что мозаичное чередование растительности и обнаженной земли (куда саранча откладывает яйца) благоприятствует экспоненциальному росту численности популяции. По-видимому, мы имеем здесь дело с популяционным взрывом, обусловленным и нестабильностью окружающей среды, и простотой условий. Вероятно, как и у леммингов, не каждый популяционный максимум сопровождается эмиграцией, поэтому частота повторения этих бедствий не обязательно отражает точную периодичность колебаний плотности. И даже при этом за период между 1695 и 1895 гг. вспышки численности регистрировались по крайней мере каждые 40 лет. Более современный исторический обзор, посвященный подъемам и спадам численности пустынной саранчи, принадлежит Валову (Waloff, 1966).
* (Одиночные и мигрирующие формы обнаружены у нескольких видов саранчи; их часто описывали как разные виды, пока не были установлены их истинные взаимоотношения.)
Известен интересный пример колебаний численности типа хищник - жертва с участием и животных, и растений. Продукция семян у хвойных часто циклична, и соответствующие колебания часто наблюдаются у зерноядных птиц и других животных.
Регулярные циклы численности, такие, как у рыси и зайца или у леммингов, представляются парадоксальными и загадочными: ведь в природе изменения часто далеко не столь регулярны. Эти циклы имеют две поразительные особенности: 1) сильней всего они выражены в менее сложных экосистемах северных районов, как явствует из приведенных примеров; и 2) хотя пики численности могут одновременно отмечаться на обширных пространствах, у одного вида, обитающего в разных районах, ее пики отнюдь не всегда совпадают по времени. Теории, выдвинутые для объяснения этих регулярных циклов, относятся ко всем уровням иерархической организации, и их можно подразделить на: 1) метеорологические теории; 2) теории случайных флуктуаций; 3) теории взаимодействия популяций и 4) теории взаимодействия трофических уровней. Ниже будут даны краткие характеристики каждой из этих теорий.
Все попытки связать регулярные колебания численности а климатическими факторами остаются пока безуспешными, несмотря на то, что синхронность и четкая выраженность этих колебаний в северных районах, казалось бы, свидетельствуют о периодических событиях за пределами локальной экосистемы. Одно время полагали, что циклы численности рыси и другие 10-летние циклы объясняются цикличностью появления солнечных пятен, с которым связаны основные изменения погоды. Однако, Мак-Лалич (MacLulich, 1937) и другие показали, что на самом деле корреляция здесь отсутствует. До сих пор не выявлено широкораспространенной климатической периодичности с 3-4-летними интервалами.
Палмгрен (Palmgren, 1949) и Коул (Cole, 1951, 1954) высказали предположение, что колебания, кажущиеся нам регулярными, возможно, являются следствием случайных изменений комплекса биотических и абиотических условий среды, в которых, обитает популяция. Если эта гипотеза верна, то ни один из факторов нельзя считать важнее остальных. Однако Кейт (Keith, 1963), проведя детальный статистический анализ циклов северных млекопитающих и птиц, пришел к выводу, что десятилетние циклы неслучайны, хотя доказать, что более короткие циклы не являются следствием случайных флуктуаций, все же трудно.
Если будет доказано, что случайные и иные климатические факторы не являются основной причиной резких колебаний численности, то вполне естественно искать причины этих колебаний в самих популяциях ("внутренние факторы"). Имеются некоторые данные, свидетельствующие о том, что, по-видимому, существуют какие-то механизмы, которые могут действовать сопряженно с изменениями погоды или других физических факторов.
Основываясь на медицинской теории стресса (общего адаптационного синдрома), созданной Гансом Селье (Hans Selye), Кристиан и его сотрудники (см. Christian, 1950, 1961, 1963; Christian, Davis, 1964) собрали многочисленные данные как для природных, так и для лабораторных популяций, показывающие, что у высших позвоночных в условиях перенаселенности происходит увеличение надпочечников. Это один из симптомов сдвига нейроэндокринного равновесия, который в свою очередь сказывается на поведении животных, их репродуктивном потенциале, устойчивости к болезням и другим видам стресса. Комплекс таких изменений часто бывает причиной резкого снижения плотности популяции. Например, зайцы-беляки в периоды пика численности часто внезапно погибают от "шоковой болезни", которая, как было показано, связана с увеличением надпочечников и другими признаками нарушения гормонального равновесия.
Читти (Chitty, 1960, 1967) высказал предположение, что различия в агрессивном поведении и выживаемости, наблюдающиеся в различные фазы цикла у полевок, обусловлены генетическими сдвигами - ситуация, сходная с той, которая отмечена у коконопряда. Таким образом, совершенно очевидно, что адаптационный синдром сглаживает колебания.
Теории четвертой группы основываются на представлении, согласно которому регуляция циклов численности осуществляется на экосистемном, а не на популяционном уровне. Разумеется, изменения, которые охватывают несколько порядков величин, должны распространяться не только на взаимодействия высших трофических уровней, например на взаимодействие хищник - жертва, но и на взаимодействия, характерные для первичного звена пищевой цепи, такого, как растения - травоядные. Примером служит описанная ранее гипотеза Холлинга, основанная на исследованиях листовертки-почкоеда. Другим примером служит предложенная для объяснения циклов численности полевок в тундре гипотеза "возврата биогенных элементов" (Schultz, 1964, 1969; Pitelka, 1964, 1973). Согласно этой гипотезе, подтвержденной данными изучения круговоротов биогенных элементов, в результате сильного выедания растительности леммингами в годы пика их численности в последующие годы происходит связывание и уменьшение доступности биогенных элементов (особенно фосфора), в результате чего питательная ценность пищи леммингов сильно снижается. В связи с этим рост и выживание молодых особей ограничиваются. На протяжении третьего и четвертого года биогенные элементы возвращаются в круговорот, растительный покров восстанавливается, и экосистема опять может поддерживать высокую плотность леммингов.
Мы уделили столь большое внимание случаям резкого изменения численности не потому, что они так уж часто встречаются в природе, а потому, что при их исследовании выявляются функции и взаимодействия, вероятно, имеющие более общий характер, но не всегда с очевидностью проявляющиеся в тех популяциях, где плотность меняется менее резко. Проблема циклических колебаний в каждом конкретном случае может быть сведена к установлению того, определяются ли они в основном одним или несколькими факторами или же их причины столь многочисленны, что разобраться в них не представляется возможным, хотя и ясно, что общая структура взаимосвязей может быть понята на основе того, что Коул (Cole, 1957) назвал "вторичной простотой", имея в виду, что закономерностей здесь "не больше, чем встречается в последовательности случайных чисел". Первая ситуация, конечно, возможна в простых экосистемах, как экспериментальных, так и природных; последняя более вероятна для сложных экосистем.
Рассмотрев несколько интересных конкретных случаев, перейдем к анализу более общей проблемы регуляции численности популяций.
При использовании материалов сайта активная ссылка обязательна:
http://ecologylib.ru/ 'Зелёная планета - экология и охрана природы'